

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
A class of pattern recognition receptors (PRRs), toll-like receptors (TLRs), is expressed in immune cells (e.g., monocytes/macrophages, dendritic cells, T cells, and B cells) as well as non-immune cells, such as epithelial cells. In total, 10 TLRs have been identified in humans and 12 in mice (1). Each TLR recognizes a specific molecular pattern of pathogen called pathogen-associated molecular patterns, such as lipoprotein (TLR2 agonist), dsRNA (TLR3 agonist), lipopolysaccharide (LPS, TLR4 agonist), flagellin (TLR5 agonist), ssRNA (TLR7/8 agonist), and unmethylated CpG DNA (TLR9 agonist) (2). In innate immune cells, TLR signaling induces the production of various pro-inflammatory cytokines leading to innate immune responses (34). Most TLRs act as homodimers, but TLR2 functions as a heterodimer with either TLR1 or TLR6. TLR1/2 heterodimer senses triacyl lipoproteins, whereas TLR2/6 heterodimer senses diacyl lipoprotein (5). TLR2 is important in recognizing gram-positive bacteria, whereas TLR4 recognizes gram-negative bacteria in innate immune response (6).
B cell receptor (BCR) cross-linking with Ag, B cell–T cell (CD40–CD40L) interaction, and the optimal TLR stimulation of B cells is sufficient for B cell activation, proliferation, and plasma cells expressing high-affinity Abs. Cytokines produced by other immune cells, such as T cells and macrophages, directly help B cells to activate and undergo Ig class switching. Ig class switch recombination (CSR) is a mechanism that causes the production of Abs to change from IgM or IgD to other Ab isotypes, namely, IgG, IgE, or IgA, which have defined roles in the humoral immunity of the immune system. Ig CSR occurs between switch regions located 5′ to each constant region gene of IgH (CH) except for δ. The transcription of the corresponding unrearranged CH gene produces germline transcripts (GLT), which are prerequisites for subsequent Ig CSR (78). Activation-induced cytidine deaminase (AID) is also essential for the process, Ig CSR (9). T cell-dependent B cell Ab responses produce high-affinity Abs for antigens but require approximately 1 week, which a rather long delay for defense against fast-replicating pathogens such as pathogenic Escherichia coli and influenza virus. To overcome this limitation, extrafollicular B cells rapidly undergo Ig CSR, seemingly through TLR-mediated T cell-independent pathway (10). As a result, class switched-IgG, IgE, and IgA as well as IgM are produced. Consistent with the concept, naïve B cells proliferate and secrete Abs to various TLR agonists in vitro, in the absence of BCR cross-linking or T cell help (11).
TLR stimulation has been reported to affect B cell functions directly without support by T cells (12). In mouse, TLR4 agonist LPS promotes B cell proliferation and differentiation into plasma cells (1314). In addition, LPS induces IgG2b and IgG3 class switching (1516). TLR9 agonist CpG also strongly induces B cell proliferation and differentiation (1217). Furthermore, CpG inhibits LPS/IL-4 induced IgG1 and IgE production, while it promotes IgG2a, IgG2b, and IgG3 production (1819). IgG2a production induced by CpG is reported to result directly from induction of the transcription factor T-bet, which is important to induce GLTγ2a expression (1820). The suppression of IgG1 and IgE production by CpG may be mediated by the inhibition of NF-κB and IRF4 activity (21). TLR7/8 agonist R-848 directly inhibits anti-CD40/IL-4-mediated IgE production (22). Several reports have shown that TLR signaling interplays with other receptor signaling such as other TLRs, nod-like receptors (e.g., Nod1 and Nod2), and BCR (2324). LPS promotes B cell maturation, but this is arrested upon TLR2 stimulation (25). In addition, BCR signaling synergizes with TLR signaling for AID expression and Ig CSR (26). Furthermore, in humans, Nod1 and Nod2 cooperate with BCR and TLR, but not anti-CD40, to induce B cell activation (27). Recently, we found that the C-type lectin receptor Dectin-1 selectively induces IgG1 class switching by LPS-activated mouse B cells (28).
Thus, many studies have focused on the direct effect of stimulation of B cell PRRs (i.e., TLRs, nod-like receptors, C-type lectin receptor) on B cell activation and Ig class switching. Nevertheless, the role of TLR1/2 on B cells in activation and differentiation of B cells is not clear. Hence, in this study, we used purified untouched mouse spleen resting B cells and investigated the direct effect of stimulation of the TLR1/2 agonist Pam3CSK4 on B cell viability, proliferation, activation, isotype switching, and Ig production in the absence or presence of TLR4 agonist LPS.
MATERIALS AND METHODS
Animals
BALB/C mice were purchased from Damool Science (Daejeon, Korea) and maintained on an 8:16 h light/dark cycle in an animal environmental control chamber. Eight- to twelve-week-old mice were used, and animal care was conducted in accordance with the institutional guidelines of the Institutional Animal Care and Use Committee of Konyang University.
Cell culture and reagents
The mouse B lymphoma cell line CH12F3-2A (surface µ+) (29) was provided by Dr. T. Honjo (Kyoto University, Kyoto, Japan). The mouse B cell lines, L10A6.2 (surface µ+, mature B cell line) and A20.3 (surface γ2a+), were provided by Dr. J. Stavnezer (University of Massachusetts Medical School, Worcester, MA, USA). Untouched mouse spleen resting B cells were obtained by depletion of CD43+ cells using anti-CD43 microbeads and high-gradient magnetic cell separation according to manufacturer's instruction (MACS; Miltenyi Biotec, Bergisch Gladbach, Germany). Briefly, BALB/c spleen cell suspensions were washed with Hank's balanced salt solution (HBSS; WelGENE, Daegu, Korea) and treated with 0.83% ammonium chloride to lyse red blood cells (RBCs). The spleen cells were treated with anti-mouse CD43 microbeads and depleted with LS column and MACS Separator (Miltenyi Biotec). The purity of resting B cells (CD43-B220+) assessed by FACSCalibur (BD Biosciences, San Jose, CA, USA) following staining of the cells with anti-CD43 FITC (eBioscience, San Diego, CA, USA) and anti-B220 PE (BD Biosciences). The cells were cultured at 37°C in a humidified CO2 incubator (Forma Scientific, Marietta, OH, USA) in RPMI-1640 medium (WelGENE) supplemented with 10% fetal bovine serum (PAA Laboratories, Etobicoke, ON, Canada). The cells were stimulated with Pam3CSK4 (Pam3CysSerLys4: synthetic bacterial tripalmitoylated lipopeptide - TLR1/2 ligand; Invivogen, San Diego, CA, USA) and LPS (ultra pure LPS, E. coli 0111:B4; Invivogen). The mouse macrophage cell line RAW264.7 was cultured in DMEM (WelGENE, 2 mM L-glutamine; 100 U/ml penicillin; 100 μg/ml streptomycin) plus 10% fetal bovine serum in a humidified CO2 incubator.
Cell viability, proliferation, and activation assays
Cell viability was determined by either trypan blue exclusion test or EZ-Cytox cell viability assay (DaeilLab Service Co., Ltd., Seoul, Korea) according to manufacturer's instructions (28). For cell proliferation assay, purified mouse resting B cells were labeled with CFSE (eBioscience) and then added with Pam3CSK4 and LPS. CFSE dilution was measured by counting 10,000 cells with a FACSCalibur. For cell activation assay, cultured cells were stained with anti-CD69-FITC (BD Biosciences) and the expression levels were analyzed by flow cytometry (FACSCalibur).
Isotype-specific ELISA
Abs produced in B cell cultures were detected using isotype-specific ELISAs as previously described (28).
RT-PCR
RNA preparation and RT-PCR were performed as previously described (28). The PCR primers [for TLRs (30); for GLTs (31); for AID (32); for T-bet (designed by Primer3 software); for Blimp-1, XBP-1, IRF-4, Pax5, BCL6, and c-myc (33); for ELL2 (34)] were synthesized by Bioneer (Daejeon, Korea): TLR1, forward 5′-GGACTTCCACATGTCTCCACTATCC-3′, reverse 5′-TCCATGC TTGTTCTTCTCTGTGG-3′, (product size, 569 bp); TLR2, forward 5′-GTGGTACC TGAGAATGATGTGGG-3′, reverse 5′-TTAAGGAAGTCAGGAACTGGGTG-3′, (product size, 541 bp); TLR4, forward 5′-CTGGGTGAGAAATGAGCTGG-3′, reverse 5′-GATACAATTCCACCTGCTGCC-3′, (product size, 249 bp); GLTγ1, forward 5′-CAGCCTGGTGTCAACTAG-3′, reverse 5′-CTGTACATATGCAAGGCT-3′ (product size, 532 bp); GLTε, forward 5′-ACTAGAGATTCACAACG-3′, reverse 5′-AGCGATGAATGGAGTAGC-3′ (product size, 423 bp); GLTγ2a, forward 5′-GCTGATGTACCTACCTGAGAGA-3′, reverse 5′-GCTGGGCCAGGTGCTCGAGGTT-3′, (product size, 394 bp); GLTγ2b, forward 5′-GGGAGAGCACTGGGCCTT-3′, reverse 5′-AGTCACTGACTCAGGGAA-3′ (product size, 318 bp); GLTγ3, forward 5′-CAAGTGGATCTGAACACA-3′, reverse 5′-GGCTCCATAGTTCCATT-3′ (product size, 349 bp); GLTα, forward 5′-CTACCATAGGGAAGATAGCCT-3′, reverse 5′-TAATCGTGAATCAGGCAG-3′ (product size, 206 bp); AID, forward 5′-AGATAGTGCCACCTCCTGCTCACTGG-3′, reverse 5′-GGCTGAGGTTAGGGTTCCATCTCAG-3′ (product size, 209 bp); T-bet, forward 5′-GTCGCTTCCTTGGATCCTTC-3′, reverse 5′-TCCACCAAGACCACATCCAC-3′ (product size, 373 bp); Blimp-1, forward 5′-CCCGCGGCCGTAGAAAA-3′, reverse 5′-GGATGCCTCGGCTTGAACAG-3′ (product size, 350 bp); XBP-1, forward 5′-GCTGGAGCAGCAAGTGGTGGATTTGG-3′, reverse 5′-GGCTTCCAGCTTGGCTGATGAGGTCC-3′ (product size, 418 bp); IRF-4, forward 5′-GGACTACAATCGTGAGGAGGAC-3′, reverse 5′-ACGTCACAGGACATTGATATGG -3′ (product size, 413 bp); Pax5, forward 5′-ACCGCGTGTTTGAGAGACAG-3′, reverse 5′-TTGGGGAACCTCCAAGAATC-3′ (product size, 373 bp); BCL-6, forward 5′-CAGCACCTTCCTCTTCTCTGATGAGGAGCTCC-3′, reverse 5′-CTGGCGGAGAGCCAGAGGCCTGAAGGATGC-3′ (product size, 485 bp); c-myc, forward 5′-CTCCGGGCTCTGCTCTCCATCCT-3′, reverse 5′-GGGGGTGCGGCGTAGTTGTGC-3′ (product size, 741 bp); ELL2, forward 5′-GAGAGGAAAAGGTCAACGCC-3′, reverse 5′-GGCTGGTGCAGCATTTGA-3′ (product size, 367 bp); and β-actin, forward 5′-CATGTTTGAGACCTTCAACACCCC-3′, reverse 5′-GCCATCTCCTGCTCGAAGTCTAG-3′ (product size, 318 bp). cDNA synthesis kit and PCR reagents were purchased from NanoHelix (Daejeon, Korea) and iNtRON Biotechnology (Seongnam, Korea), respectively. PCR for β-actin were performed in parallel to normalize cDNA concentrations within each set of samples. PCR products were resolved by electrophoresis on 2% agarose gels.
Cell surface analysis to detect plasma cells
The purified resting B cells were stimulated for 3 or 4 days and then collected. The cells were stained with rat anti-mouse CD138 PE (BD Pharmingen, San Diego, CA, USA) and rat anti-mouse CD45R/B220 FITC (BD Pharmingen). The proportion of plasma cells (CD138+B220lo) was assessed by flow cytometric analysis with a FACSCalibur.
RESULTS
Effect of Pam3CSK4 on mouse B cell growth and Ig production
We used untouched resting B cells (CD43−B220+) purified from mouse spleen to investigate the direct effect of TLR1/2 agonist Pam3CSK4 on B cell activation and differentiation. The purity of resting B cells assessed by flow cytometric analysis was higher than 98% (Fig. 1A). Many studies have reported that mouse and human B cells express various TLRs (122435363738). First, we verified whether the purified resting B cells express TLR1, TLR2, and TLR4. As shown in Fig. 1B, resting B cells as well as 3 mouse B cell lines (CH12F3-2A, L10A6.2, and A20.3) expressed TLR1, TLR2, and TLR4 mRNAs (Fig. 1B). RAW264.7 mouse macrophage cell line was used as a positive control for the TLRs expression (24). We stimulated resting B cells with 0.5 µg/ml of Pam3CSK4 or 1 µg/ml of LPS, a well-known potent mouse B cell mitogen (Fig. 1C), to examine the effect of Pam3CSK4 on B cell proliferation, viability, and Ig production. Pam3CSK4 highly increased cell proliferation and sustained cell viability up to 3 days of culture. LPS increased cell proliferation much more slowly than Pam3CSK4, but steadily sustained cell viability up to 4 days. At least in part, Pam3CSK4 increased cell proliferation and sustained cell viability during overall culture time, and this ability is much better than LPS. Thus, Pam3CSK4 can act as a potent mitogen for mouse B cells. Then, we determined Ig production by Pam3CSK4- or LPS-stimulated B cells in the same culture. The levels of IgM, IgG2b, and IgG3 produced by Pam3CSK4-stimulated B cells were not quite different from LPS stimulation, whereas Pam3CSK4 much more enhanced IgG2a and IgA production compared with LPS (Fig. 1D). Interestingly enough, we observed that Pam3CSK4 only barely induced IgG1 secretion by B cells even though its enhancing effect on cell growth was better. We could not detect any IgE production in this culture (data not shown). These results indicate that Pam3CSK4 is not an appropriate TLR agonist to steadily induce IgG1 production by B cells, although it is a good candidate for B cell mitogen.
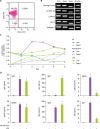 | Figure 1Effects of Pam3CSK4 and LPS on mouse B cell viability and Ig production. (A) Purity of isolated mouse spleen resting B cells (CD43−B220+) was measured by flow cytometric analysis. Data shown are representative of all the experiments carried out in this study. (B) Expression of TLR1, TLR2, and TLR4 in the isolated mouse spleen resting B cells (CD43−B220+). RNAs were isolated from the resting B cells and the indicated cell lines, and the levels of TLR1, TLR2, and TLR4 mRNA were measured by RT-PCR. (C) Effects of Pam3CSK4 and LPS on mouse B cell viability. The resting B cells (1.5×106 cells/ml) were cultured with Pam3CSK4 (0.5 µg/ml) or LPS (1 µg/ml). Cell viability was assessed using trypan blue exclusion test at the indicated days. Data shown are representative of 2 independent experiments. (D) Effects of Pam3CSK4 and LPS on Ig production. Culture conditions were the same as in (C). After 7 days of culture, supernatants were harvested and the levels of Ig production were determined by isotype-specific ELISA. Data shown are averages of quadruplicate cultures with SEM error bars.
SEM, standard error of the mean.
*p<0.05, **p<0.01.
|
Pam3CSK4 increases B cell growth but decreases IgG1 production in a dose-dependent manner
To further elucidate the role of Pam3CSK4 in B cell growth and Ig production, we stimulated resting B cells with various doses of Pam3CSK4 and measured cell proliferation, cell viability, and Ig production. Pam3CSK4 dose-dependently increased both cell proliferation (Fig. 2A) and cell viability (Fig. 2B), which were maximal at about 1 µg/ml of Pam3CSK4 (Fig. 2C). These data indicate that B cell TLR1/2 saturation with Pam3CSK4 was achieved at the agonist concentrations higher than 1 µg/ml. On the other hand, Pam3CSK4 increased the productions of IgM, IgG2a, IgG2b, IgG3, and IgA, while IgG1 production was selectively decreased in a dose-dependent manner (at >0.25 µg/ml) (Fig. 2D). These results suggest that excess Pam3CSK4 specifically has less effect on IgG1 production by mouse B cells, although it is a potent B cell mitogen and a stimulant to enhance other Ig isotype production.
 | Figure 2Dosage effect of Pam3CSK4 on B cell proliferation, viability, and Ig production. Mouse spleen resting B cells were stimulated with the indicated concentrations of Pam3CSK4. After 2 and 3 days of culture, cell proliferation was measured by CFSE assay (A) and cell viability was measured by EZ-Cytox assay (B). (C) Overlap graph of cell proliferation (A) and viability (B) at 3 days of culture. Low CFSE intensity cells (%) means the proportion of proliferating cells. (D) After 7 days of culture, supernatants were harvested and the levels of Ig production were determined by isotype-specific ELISA. Data shown are representative of 2 independent experiments.
|
Combination effect of Pam3CSK4 and LPS on B cell proliferation, viability, activation, and Ig production
Different TLR agonists can result in a synergistic effect on biological activities including cytokine production by immune cells. TLR4 signaling promotes B cell maturation, while TLR2 inhibits that process (25). Therefore, in this study, we further investigated the roles of TLR1/2 agonist Pam3CSK4 and TLR4 agonist LPS in B cell proliferation, viability, and Ig production. TLR4 agonist LPS is a mouse B cell mitogen and polyclonal activator and induces switching to IgG3 and IgG2b (1516). As shown in Supplementary Fig. 1, LPS dose-dependently increased B cell growth (proliferation and viability) and all Ig production. To determine whether 2 different TLR agonists, Pam3CSK4 and LPS, have synergistic/antagonistic effect on B cell function, we stimulated resting B cells with 0.5 µg/ml of Pam3CSK4 and/or 5 µg/ml of LPS. As shown in Fig. 3A and B, Pam3CSK4 further enhanced not only LPS-induced cell proliferation but also cell viability. We examined the expression of surface CD69, a lymphocyte activation marker, on LPS and/or Pam3CSK4-stimulated B cells to evaluate the effect of Pam3CSK4 and LPS on B cell activation. LPS and Pam3CSK4 induced B cell activation (approximately 28% increase and 32% increase, respectively), and this increase was further enhanced by their combination (approximately 54% increase) (Fig. 3C). Pam3CSK4 did not affect LPS-induced IgM and IgG2b secretion by B cells, whereas it dramatically decreased LPS-induced IgG1 and IgG3 production (Fig. 3D). In addition, Pam3CSK4-induced IgA production was decreased by LPS. By contrast, LPS-induced IgG2a production was further reinforced by Pam3CSK4. In addition, we observed that Pam3CSK4 enhances LPS-induced cell growth and regulates each Ig production in a dose-dependent manner (Supplementary Fig. 2). These results indicate that TLR1/2 agonist Pam3CSK4 and TLR4 agonist LPS play diverse roles in each Ig production, even if both are potent B cell activator and they synergistically induce B cell growth and activation.
 | Figure 3Combination effect of LPS and Pam3CSK4 on B cell proliferation, viability, activation, and Ig production. The resting B cells were stimulated with LPS (5 µg/ml) and Pam3CSK4 (0.5 µg/ml). After 2 and 3 days of culture, cell proliferation was measured by CFSE assay (A), and cell viability was measured by EZ-Cytox assay (B). (C) After 2 days of culture, cells were stained with anti-CD69-FITC and surface CD69 expression was analyzed by flow cytometry. (D) After 7 days of culture, supernatants were harvested and the levels of Igs production were determined by isotype-specific ELISA. Data shown are averages of quadruplicate cultures with SEM error bars.
SEM, standard error of the mean; NS, not significant.
*p<0.05, **p<0.01, ***p<0.001.
|
Effects of Pam3CSK4 and LPS on the expression of GLT
To elucidate the mechanism of differential regulation of Ig production by Pam3CSK4 and LPS, we analyzed the expression of each GLT, which is a prerequisite for each Ig CSR, by Pam3CSK4 and/or LPS-stimulated mouse resting B cells (Fig. 4). Pam3CSK4 alone and LPS alone induced all GLT expression. Interestingly, Pam3CSK4 diminished LPS-induced GLTγ1 and GLTε expressions, whereas it enhanced LPS-induced GLTγ2a. However, Pam3CSK4 did not affect LPS-induced GLTγ2b, GLTγ3, and AID expression. We could not detect any GLTα expression in this culture (data not shown). Then, we tested the kinetic effect of Pam3CSK on LPS-induced GLTγ1 and GLTγ2a expression. As shown in Fig. 5, Pam3CSK4 consistently decreased LPS-induced GLTγ1 expression from day 3 to day 5, whereas it enhanced LPS-induced GLTγ2a from day 1 to day 3. Expression pattern of GLTε was similar to those of GLTγ1. Here Pam3CSK4 additively stimulated LPS-induced cell growth and differentially regulated each Ig production as described in Fig. 3D (Supplementary Fig. 3). Transcription factor T-bet is required for the induction of GLTγ2a transcription, resulting in IgG2a CSR (39). T-bet mRNA expression was increased when Pam3CSK4 and LPS were treated in combination at days 2 and 3 (Fig. 5). Collectively, these results indicate that Pam3CSK4, at least in part, regulates LPS-induced IgG1 and IgG2a production through the modulation of their GLTs, i.e., GLTγ1 and GLTγ2a, respectively.
 | Figure 4Effect of Pam3CSK4 on LPS-driven GLT expression. The resting B cells were stimulated with LPS (5 µg/ml) and Pam3CSK4 (0.5 µg/ml). After 3 days of culture, RNAs were isolated, and the levels of GLTs and AID mRNA were measured by RT-PCR (upper panel). cDNA from each sample were prepared at 1/5 and 1/25 dilutions, and the expression levels of GLTs were measured by PCR (lower panel). Data were obtained from 2 independent experiments (Exp1 and Exp2).
|
 | Figure 5Kinetic effect of LPS and Pam3CSK4 on GLT expression. The resting B cells were stimulated with LPS (5 µg/ml) and Pam3CSK4 (0.5 µg/ml). After the indicated days of culture, RNAs were isolated and the levels of GLTs and T-bet mRNA were measured by RT-PCR. Graphs show relative GLTγ1 and GLTγ2a cDNA levels normalized to the expression of β-actin cDNA by ImageJ (National Institutes of Health, Bethesda, MD, USA) analysis. Densitometric data are averages of 2 independent experiments with ranges (bars).
|
Effects of Pam3CSK4 and LPS on plasma cell differentiation
To test whether the pattern of differential Ig production by Pam3CSK4 is due to the regulation of plasma cell differentiation in the stimulated B cells, we analyzed mRNA expression pattern of various transcription factors (3340), which are related to plasma cell differentiation. Blimp-1 is a key molecule in plasma cell development and directly represses Pax5 expression. Pax5 is required for the maintenance of B cell identity and represses the expression of XBP-1, itself an essential player in plasma cell development. IRF4 is an essential transcriptional regulator of Ab-secreting cell function. Repression of c-myc by Blimp-1 is necessary for terminal differentiation of B cells. BCL6 is also a B cell maintenance factor and acts as a repressor of Blimp-1. ELL2 is essential for the shifting of Igs from a membrane form to a secrete form in plasma cells (34). Both LPS and Pam3CSK4, respectively, increased Blimp-1, XBP-1, IRF-4, and ELL2 expression, whereas there was no any combination effect of Pam3CSK4 and LPS on those expressions (Fig. 6A). In addition, Pax5, BCL6, and c-myc expressions were not changed in the condition of Pam3CSK4 in combination with LPS. Furthermore, Pam3CSK4- or LPS-induced CD138hiB220lo plasma cell population was not altered when they were stimulated in the combination (Fig. 6B, C). Overall, these results suggest that TLR1/2 and TLR4 stimulation induces plasma differentiation, but there is neither a synergistic nor an antagonistic effect between those TLRs on it.
 | Figure 6Effects of LPS and Pam3CSK4 on plasma cell differentiation. Resting B cells were stimulated with LPS (5 µg/ml) and Pam3CSK4 (0.5 µg/ml). (A) After 2.5 days of culture, RNAs were isolated and the levels of indicated mRNAs measured by RT-PCR. (B, C) Proportions of CD138+ (Syndecan-1+) B220lo plasma cells. After 3 and 4 days of culture, harvested cells were stained with anti-CD138 PE and anti-B220 FITC Abs and analyzed by FACSCalibur. (B) Data shown are representative (3 days culture) of 2 independent experiments. (C) Data are averages of 3 and 4 days cultures from 2 independent experiments with SEM error bars.
SEM, standard error of the mean.
|
DISCUSSION
In the present study, we demonstrated that TLR1/2 agonist Pam3CSK4 acts as a potent mouse B cell mitogen (an activator) commonly with TLR4 agonist LPS, and Pam3CSK4 differentially regulates LPS-induced Ig (especially IgG1 and IgG2a) production through the modulation of the GLT expression (GLTγ1 and GLTγ2a, respectively).
Pam3CSK4 selectively diminished the expression of LPS-induced GLTγ1/ε, whereas it enhanced GLTγ2a (Fig. 4). IL-4 and IFN-γ induce IgG1 and IgG2a production, respectively, by mouse B cells stimulated with LPS (4142). To determine the autocrine effect of IL-4 and IFN-γ on GLT expression in the same culture, we examined the levels of IL-4 and IFN-γ production from the Pam3CSK4- and/or LPS-stimulated mouse B cells. We did not detect any IL-4 and IFN-γ production in the culture (data not shown). IL-12 was also known to negatively regulate IgG1 and IgE syntheses (4344), and IL-12 and CpG synergistically induces T-bet expression and IgG2a production by mouse B cells (18). Pam3CSK4 was reported to enhance IL-12 production by dendritic cells in vitro (45). IL-12 is a heterodimeric protein produced by B cells, phagocytic cells, and other antigen-presenting cells (46). Both human and mouse B cells produce large amounts of IL-12 in response to combined stimulation with BCR, CD40 and CpG (1847). LPS has been shown to stimulate IL-12 production in host cells and consequently down-regulate Th2 responses in animal models of allergic airway inflammation. Thus, it is possible that Pam3CSK4 and LPS stimulate mouse B cells to produce IL-12, which is involved in regulation of IgG1/IgE and IgG2a production. Further studies will examine this possibility.
Other possibilities will explain the inhibiting or enhancing effect of Pam3CSK4 on LPS-induced GLTγ1/ε or GLTγ2a transcription. First, Pam3CSK4 may block LPS-mediated GLTγ1 expression through NF-κB pathway. Previous studies reported that LPS induces NF-κB complexes p50-cRel and p50-p50 dimers, while CD40L induces p50-p65 and p50-RelB dimers in spleen B cells (4849). LPS-induced p50-cRel/p50-p50 complex inhibits CD40L-induced p50-p65/p50-RelB complex by competitively binding to GLTγ1 promoter, resulting in diminishing GLTγ1 promoter activity (48). In addition, CTLA-4 was reported to decrease GLTγ1 and GLTε expression by inhibiting NF-κB activation, resulting in inhibition of IgG1 and IgE production (50). This indicates that different induction of NF-κB/Rel proteins has different effect on GLTγ1 transcription. NF-κB subunit p65 is activated by Pam3CSK4, which is important for Pam3CSK4-medicated B cell activation (5152). Therefore, it is possible that LPS-induced NF-κB activation for GLTγ1 transcription was blocked by either binding of other Pam3CSK4-induced NF-κB complexes on GLTγ1 promoter region or competition between LPS- and Pam3CSK4-induced NF-κB subunits for functional NF-κB complex formation. On the other hand, previous report demonstrated that phosphoinositide 3-kinase (PI3K)-mediated signaling negatively regulates LPS responses in macrophages (53). Furthermore, the cytosolic domain of TLR2 was reported to contain PI3K binding motif, and TLR2 activates PI3K-mediated signaling (51). Therefore, Pam3CSK4 may directly inhibit LPS-induced GLTγ1 expression via the PI3K pathway (5455). In addition, previous reports demonstrated that transcription factors BCL6 and ID2 repress IgE CSR (555657). Indeed, we are currently exploring these possibilities whether Pam3CSK4-induced/activated transcription factors, such as NF-κB, BCL6, and ID2, repress LPS-induced GLTγ1/ε promoter activity and which signal pathways were involved.
We showed that transcription factor T-bet expression was further enhanced by Pam3CSK4 in combination with LPS (Fig. 5). T-bet acts as a transcription factor on GLTγ2a promoter to induce class switching to IgG2a in B cells (2039). Indeed, TLR9 agonist CpG directly induces T-bet expression in B cells, resulting in increasing IgG2a (“Th1-like” Ig) production, and inhibits IgG1 and IgE (“Th2-like” Igs) class switching in B cells (1819585960). In addition, CpG suppresses IgE CSR by reducing NF-κB binding to the GLTε promoter, but not through transcription factor ID2 or BCL6 (21). STAT1 is also a key transcription factor to activate IFNγ-induced GLTγ2a transcription. Pam3CSK4 did not further enhance LPS-induced STAT1 mRNA expression (data not shown). However, whether Pam3CSK4 affects STAT1 activation (phosphorylation) in combination with LPS stimulation in mouse B cells remains to be tested. Thus, we speculate that Pam3CSK4 negatively and positively regulates LPS-induced GLTγ1/ε and GLTγ2a expression, respectively, by enhancing T-bet expression, resulting in the regulation of LPS-induced IgG1/IgE and IgG2a production.
We observed that, although Pam3CSK4 inhibits LPS-induced IgG3 production (Fig. 3D), LPS-induced GLTγ3 expression was not altered by Pam3CSK4 treatment (Fig. 4). IFN-γ inhibits LPS-mediated switching to IgG3 (41). Interestingly, IFN-γ suppression of LPS-induced switching to IgG3 is not associated with a corresponding reduction in the levels of GLTγ3 (6162). Therefore, Pam3CSK4 may suppress LPS-induced IgG3 production through other pathways independent of the GLTγ3.
In this study, we used Pam3CSK4, which is a ligand for the TLR1/2 complex. TLR2 forms with either TLR1 or TLR6 and is recognized differently according to the type of complexes. Therefore, to further verify the role of TLR2, it is necessary to test other TLR2 agonists, such as MALP-2, Pam3Cys (TLR2/6 agonist), and PGN (TLR2 agonist). Indeed, we have tested the effect of TLR2/6 agonist Pam2CSK4 on B cell proliferation, GLT expression, and Ig production. We observed that, like Pam3CSK4, Pam2CSK4 enhances LPS-induced cell proliferation, GLTγ2a expression, and IgG2a production while decreasing GLTγ1/ε expression (data not shown). Thus, TLR2 engagement on B cells is involved in their proliferation, activation, and differential Ig class switching, especially IgG1/IgE and IgG2a.
In conclusion, our findings show that the TLR1/2 engagement with Pam3CSK4 activates B cell proliferation and differentially regulates LPS-driven Ig production through the modulation of GLT expression by B cells, down-regulation of IgG1/IgE (through suppressing GLTγ1/GLTε), and up-regulation of IgG2a (through enhancing GLTγ2a). IgG2a efficiently activates complement through binding to C1q, but IgG1 does not (63), and IgG2a is appropriate for clearing viral infections (6465). High-affinity IgE and IgG1 cause anaphylaxis (6667), and abnormal overproduction of IgE causes various allergic disorders. Recently, Weir et al. (68) demonstrated that combinations of TLR adjuvants (which consisted of 2 TLR agonists, poly I:C and Pam3CSK4) promote more effective B cell activation in vitro and can be used to augment antibody responses to vaccines in vivo. Consequently, Pam3CSK4 not only can be a candidate for effectively treating IgE-mediated allergic diseases but also may be used for B cell adjuvants to enhance IgG2a response to protect infection with fast-replicating pathogens, especially gram-negative bacteria with LPS.
 XML Download
XML Download