

 PDF
PDF Citation
Citation Print
Print


Abbreviations
Bcl
B cell CLL/lymphoma
CXCR
C-X-C chemokine receptor
Dgcr8
DiGeorge syndrome critical region 8
Fox
forkhead box protein
ICOS
inducible costimulatory
IRF
interferon-regulatory factor
miRNA
microRNA
Socs
suppressor of cytokine signaling
STAT
signal transducer and activator of transcription
TFH
follicular T helper
TFR
follicular regulatory T
Th
T helper
Treg
regulatory T cell
UTR
untranslated region
INTRODUCTION
Humoral immunity plays an important role in host defense by conferring the sophisticated antibodies required to fight against foreign targets. The production of antibodies largely depends on the generation of plasma cells that are differentiated from naïve B cells. Upon antigenic stimulation, naïve B cells begin to proliferate and undergo a cognate interaction with primed CD4+ T cells at the T–B cell border in the secondary lymphoid tissues. Some of the primed B cells are then further differentiated into extrafollicular plasmablasts. Although these plasmablasts are short-lived, they are responsible for the first wave of antigen-specific antibody production, and mainly secrete IgM-type low-affinity antibodies; these instantly secreted antibodies are very important in providing the first line of defense against invading pathogens (12). Subsequently, these low-affinity antibodies are gradually replaced with class-switched high-affinity antibodies produced by the experienced plasma cells of the germinal center to ultimately fortify host immunity.
The germinal center is a transient structure formed in the follicles of the secondary lymphoid tissues during infection or vaccination. A small number of specific antigen-primed B cells migrate toward the B cell follicle area from the T–B cell border, proliferate massively, and begin to form the germinal center. During the proliferation stage, the germinal center B cells undergo a class-switch and somatic hypermutation, and selected germinal center B cells are further differentiated into class-switched memory B cells or plasma cells. These reactions waned as the pathogens or antigens are cleared from the system, and the germinal centers also eventually disappear. The size, numbers, and duration of germinal centers must be tightly regulated for maintaining immune homeostasis (3); indeed, dysregulation of the germinal center reaction can result in many immunological disorders, including autoimmunity, immunodeficiency, and lymphoma (34).
A small CD4+ T cell population has been observed in the germinal centers, and these cells are believed to play important roles in antibody production in humans and mice (56) because they control the Ig class-switch, survival, selection, and differentiation of germinal center B cells via cytokine secretion and ligand-mediated interactions (7). Recently, this specialized subset of CD4+ T cells, termed follicular helper T (TFH) cells, has been proposed as a key regulator of the magnitude of the germinal center reaction (8). For instance, strong functional activity and/or increased numbers of TFH cells are strongly linked with autoimmune diseases such as systemic lupus erythematosus (9). By contrast, low numbers of TFH cells cause impaired humoral immunity that eventually leads to immunodeficiency (810). Therefore, a key to understanding humoral immunity regulation mechanisms might lie in elucidating the detailed mechanisms underlying the development of TFH cells.
The correct positioning of TFH cells in the B cell area is one of the most important steps in generating functional TFH cells in the germinal centers. Therefore, it is not surprising that the earliest sign of the TFH cell fate decision is upregulation of C-X-C chemokine receptor (CXCR) 5 (11), which is positively controlled by the transcriptional factor achaete scutelike 2 (Ascl2) (12). Interestingly, the CXCR5+ antigen-primed T cells are repositioned at the T–B cell border by EBI2 and its ligand 7α,25-dihydroxycholesterol, which mediates chemotactic activity (13). These pre-TFH cells then interact with CD25+ dendritic cells to confer inducible costimulator ligand (ICOSL)-inducible costimulator (ICOS)-mediated signals, which are responsible for the expression of B cell CLL/lymphoma (Bcl) 6 and sustained CXCR5 expression on pre-TFH cells (13). In addition, the CD25+ dendritic cells result in an IL-2-deprived environment that is favorable for TFH cell development, because IL-2-mediated signal transducer and activator of transcription (STAT) 5 activation can suppress TFH cell development (131415). Finally, the pre-TFH cells move across the T–B border and reside in B cell follicles where they fully differentiate into TFH cells. Surprisingly, crossing of the border does not rely on CXCR5; instead, the interaction of ICOS-ICOSL with bystander B cells is the crucial event for this step (16). In addition to the migratory control of TFH cell differentiation, networks of various transcriptions factors are also critical for the development and functions of TFH cells. The transcription factors Bcl6, interferon-regulatory factor (IRF) 4, B cell-activating transcription factor (BATF), c-Maf, Ascl2, and STAT3 have been revealed as positive drivers, while B lymphocyte induced maturation protein 1 (Blimp-1), forkhead box protein O (FoxO) 1, forkhead box protein P (FoxP) 1, Fosl2, Krüppel-like factor (KLF) 2, and STAT5 are known as repressors of the development and functions of TFH cells (17).
Because of the fundamental impact of the amount of lineage-specific transcription factors for the commitment of the fate and plasticity of CD4+ T helper (Th) cells, even subtle changes of the expression of transcription factors and other regulatory genes have large impacts on T cell fates and functions (1718). Therefore, exploration of the mechanisms that underlie the delicate control of the expression of these genes might shed new light on understanding the T cell differentiation program and related immune modulation. MicroRNAs (miRNAs), which are endogenously expressed small (up to 22 nucleotides) single-stranded RNA species, have emerged as important post-transcriptional gene regulatory factors in various types of cells and species (19). MiRNAs integrated in an RNA-induced silencing complex recognize their target mRNAs through partial sequence complementarity and then bind to the targets to suppress their translation or result in their decay; perfect base pairing to the target sequence mediates the degradation of target mRNAs. Although the miRNA-mediated suppression of gene expression is not substantial in terms of magnitude, recent research has revealed an association between the dysregulation of miRNAs and many pathological conditions, including immunological disorders (20). In the past few years, extensive studies have been conducted to identify roles of miRNAs in the regulation of the germinal center reaction. Since the miRNAs directly interact with several target mRNAs, revealing the miRNA species that regulate the germinal center reaction and their functional target genes might shed light on the underlying molecular mechanisms. In this review, we discuss the state of the current knowledge on the miRNA-mediated regulation of TFH cell differentiation for the germinal center reaction, and highlight the current topics of debate in understanding of the molecular mechanisms of miRNA-mediated gene regulation.
ROLE OF miRNAs IN TFH CELL DIFFERENTIATION
Among the thousands of miRNAs that have been identified in many species to date, only a few hundreds of these miRNA species are expressed in any given cell type (2122). CD4+ T cells are not an exception to this general pattern, as the expression profiles of miRNAs in CD4+ T cells are dramatically changed during T cell activation and effector T cell differentiation (23). For instance, the expression levels of many miRNA species are down-regulated upon T cell activation. Only a handful of miRNAs, including miR-19b, miR-106b, miR-155, and miR-146a, are expressed at higher levels in activated T cells than in naïve CD4+ T cells (2324). A comprehensive study on the roles of miRNAs in T cells was performed using mice deficient in Dicer1 or DiGeorge syndrome critical region 8 (Dgcr8), which are key enzymes of miRNA biogenesis. Dicer1 or Dgcr8-deficient T cells showed severe defects in activation, proliferation and survival; surprisingly, however, the production of functional cytokines of Th1-like interferon-gamma was not impaired in Dicer-deficient CD4+ T cells (2526). Moreover, Dgcr8-deficient CD4+ T cells are preferentially differentiated into Th1 and Th2 cells, but show impaired generation of TFH cells (27). These results strongly suggest the specific regulatory roles of miRNAs in Th differentiation together with their general roles in T cell proliferation and survival. These findings have prompted researchers to explore the more detailed molecular mechanisms of individual miRNA species on TFH cell differentiation. To date, the potential roles of miR-17–92, miR-155, and miR-146a on TFH cell differentiation and functions have been most extensively studied. Interestingly, these miRNAs show different expression patterns with respect to TFH differentiation. Although all of these miRNAs are induced upon T cell receptor stimulation, miR-155 and miR-146a sustain high expression levels during the further progression of TFH differentiation, whereas induced miR-17–92 species waned out at this stage (242829).
Six different miRNAs, miR-17, miR-18a, miR-19a, miR-20a, miR19b, and miR-92a, are encoded by a single miR-17-92 transcript, and are expressed and processed together as a cluster. Initially, miR-17–92 was identified as a negative regulator of TFH cell function through CXCR5 suppression (30). However, more careful studies with loss- or gain-of-function approaches revealed that miR-17–92 expression in CD4+ T cells is essential for a functional germinal center reaction. Upon antigen priming in T cells, miR-17–92 induces the expression of the target gene Rora, which in turn suppresses the expression of inappropriate genes for TFH cells such Ccr6, Il1r1, Ilr2, and Il22 that are positively regulated by Roar (27). In addition, miR-17–92 allows for antigen-primed CD4+ T cells to migrate further into the B cell follicles and become functional TFH cells through regulating the strength of ICOS-mediated phosphoinositide 3-kinase (PI3K) activity by targeting the negative regulators of PI3K signaling pathway Pten and Phlpp2 (28). Interestingly, fully differentiated TFH cells retain miR-17–92 expression at low levels, which suggests that tight regulation of the miR-17–92 expression level is an important regulatory mode for ensuring an appropriate germinal center reaction. Indeed, overexpression of the miR-17–92 cluster in CD4+ T cells leads to the generation of an excess number of TFH-like cells and activated B cells, ultimately leading to lymphoproliferative disease and death (28).
In contrast to miR-17–92, induced miR-155 and miR-146a expression upon T cell receptor-mediated stimuli was found to be sustained at high levels on fully differentiated TFH cells. The B cell integration cluster (bic) gene encodes miR-155, and bic-null mice show impaired T cell-dependent antibody production resulting in failure to protect against virulent Salmonella typhimurium infection, suggesting an important role of miR-155 in humoral immunity (31). miR-155-deficient T cells are activated normally but are prone to become IL-4-producing Th2 cells via the de-repression of c-Maf in vitro, which is reminiscent of the phenomenon by which miR-17–92 deficiency relieves Rora expression and causes the induction of non-TFH cell-related genes (2731). Indeed, recent studies revealed a miR-155-mediated specific role for functional TFH cell generation via targeting Peli1 and Fosl2 in CD4+ T cells (2932). Peli1 is an important regulator of c-Rel protein, a member of the NF-κB family, by means of the ubiquitination in T cells, thus protecting against T cell intrinsic autoimmunity in mice (33). In line with previous results, miR-155 deficiency was shown to give rise to a low level of c-Rel expression due to the de-repression of Peli1 during TFH cell development. Interestingly, the low level of c-Rel expression does not affect TFH cell lineage commitment but rather leads to depletion of TFH cells in the draining lymph node, mainly due to the impaired proliferation of pre-TFH cells during development (29). Fosl2 binds to Jun and compete with BATF-containing activating (AP-1) complexes for DNA binding on AP-1-IRF composite elements (AICEs), which is necessary for TFH cell generation with IRF4 recruitment. Therefore, the miR-155-mediated repression of Fosl2 is important for determining TFH cell fate commitment (32). Taken together, these results suggest that miR-155 acts as a driver of TFH cell fate commitment and as an inhibitor of Th2 cell differentiation by regulating several genes concurrently.
miR-146a shows a similar expression pattern to miR-155 during TFH cell development, which indicates that miR-146a might also play important roles in TFH cell generation and functions. However, the ablation of miR-146a results in the accumulation of both TFH cells and germinal center B cells with increased expression of ICOS on T cells, which represents a restrictive role of this miRNA on TFH cell functions (24). Interestingly, the TFH cell-driven regulation of the germinal center reaction might occur through a regulatory interaction between miR-155 and miR-146a in T cells. In 7–10-month-old miR-146a-deficient mice, T cell-driven spontaneous germinal centers are formed followed by autoantibody production in the serum, and miR-155 knockout almost completely restored this aberrant activity of miR-146a-deficient T cells to that of wild-type T cells (32). These findings indicate the opposing roles of miR-146a and miR-155 as the brake and accelerator pedals for the function of TFH cells, respectively.
IL-2 mediated STAT5 signaling attenuates TFH cell fate commitment at early stages of the differentiation program. One study demonstrated that miR-182 is induced by IL-2 and regulates the late phase of expansion by the post-transcriptional regulation of FoxO1 (34). Inactivation of FoxO1 through the ICOS-mediated signaling pathway was also shown to be an important aspect in regulation of TFH cell differentiation at late stages (35), indicating a potent role of miR-182 in TFH cell generation. However, genetic approaches of miR-182 deficiency in the context of the miR-183/96/182 cluster have revealed a dispensable role of miR-182 in generating functional TFH cells (29).
ROLE OF miRNAs IN REGULATORY TFH CELLS
FoxP3+ regulatory T cell (Treg)-mediated immune modulation is a widely accepted concept. Initially, it was considered that homogenous FoxP3+ T cells suppress several distinct subsets of effector T cells (36). However, recent growing evidence has shifted this concept toward a mechanism by which FoxP3+ Tregs employ distinct regulatory pathways to restrict the responses of different subsets of the effector T cells (3738). The function of TFH cells is also controlled by a specialized subset of Bcl6+ FoxP3+ T cells located in the germinal centers, named follicular regulatory T (TFR) cells (3940). Moreover, TFR cells have been shown to suppress the numbers of TFH cells and germinal center B cells and consequently the amount of antibody produced. TFR cells are distinguished by TFH cells according to the expression of IL-10, glucocorticoid-induced tumor necrosis factor family receptor (GITR), and cytotoxic T lymphocyte antigen (CTLA)-4. More importantly, TFR cells share features in common with TFH cells, including high expression of CXCR5, programmed cell death protein (PD)-1, and ICOS in addition to Bcl6. Beyond the location and expression profiles of TFH cell signature molecules, the differentiation pathway of TFR cells has also been co-opted from that of TFH cells (41). Therefore, it is highly possible that several of the miRNAs described above would control the germinal center reaction through regulation of TFR cell generation and function. Indeed, the miR-17–92 dosage is positively correlated with the number of antigen-specific TFR cells, but not with that of polyclonal Tregs in the peripheral lymph nodes (27). In the absence of miR-146a in T cells, the numbers of both TFR and TFH cells are increased compared to those of wild-type T cells (24). However, miR-146a−/− TFR cells may lose their suppressive capacity due to the overexpression of STAT1, which can partially account for the spontaneous development of germinal centers over time, even in the presence of a high number of TFR cells (42).
FoxP3 is responsible for the elevated expression of miR-155 in Tregs, indicating that miR-155 exerts certain important roles in FoxP3+ Tregs (4344). Indeed, miR-155 has been shown to be responsible for maintaining the Treg pool in competitive settings via suppressing suppressor of cytokine signaling (Socs) 1 (44). Downregulation of Socs1 expression confers Tregs with survival and proliferation ability because of the enhanced STAT5 activity through IL-2. IL-2-mediated STAT5 activation is a well-known inhibitory axis for TFH cell generation (1415). Therefore, it is possible that miR-155 may suppress the generation of TFR cells from FoxP3+ Tregs to consequently enable adequate germinal center reactions. However, there has been no detailed study on the roles of miR-155 in TFR cells; thus, it would be interesting to address this issue in the future.
miR-10a is induced by transforming growth factor-beta and retinoic acid in induced Tregs, and is known to prevent the conversion of Tregs to TFH cells in Peyer's patches (45). Since miR-10a is also expressed in naturally occurring Tregs (46) and directly targets Bcl6 and its co-repressor, Ncor2 (45), it is plausible that miR-10a restrains the conversion of naturally occurring Tregs to Bcl6+ TFR cells. In that regard, TFR cell generation might be accompanied by a decrease in the miR-10a expression level, representing another possible regulatory axis for the germinal center reaction. Although there is no direct evidence to date, it is plausible that such regulation would occur during TFR cell generation, because TFH cells reduce the copy number of miR-10a during TFH cell differentiation (29). However, it is still worthwhile to explore the role of miR-10a in humoral immunity using T cell-specific loss-of-function or gain-of-function approaches.
Overall, it is clear that several miRNA species regulate TFR cell generation and function. However, this subject has received relatively less attention than the regulation of TFH cells, mainly because of the prevailing concept that regulatory mechanisms for TFR cells might be common with those of TFH cells. Nevertheless, the ontology of TFR cells is different from that of TFH cells (conversion from Tregs vs. differentiation from naïve T cells) and their functions are also in opposition. Therefore, there might be a specific regulatory axis for TFR cell generation and function, which is also fine-tuned by certain miRNAs. Defining these mechanisms would be an interesting and important task toward gaining a comprehensive understating of regulation of the germinal center reaction.
PERSPECTIVES
Over the past few years, extensive studies have uncovered the miRNA-mediated regulatory mechanisms of the germinal center reaction at both the cellular and molecular levels (47). These studies mainly focused on the development of TFH cells from naïve CD4+ T cells and their functions. Interestingly, most of the miRNAs identified to date, including the miR-17–92 cluster and miR-155, seem to share similar regulatory features by which they suppress non-TFH cell differentiation while concurrently facilitating the TFH cell development program (Fig. 1). This regulatory mode is fundamentally possible because even a single miRNA species is able to bind to numerous different genes and repress their expression simultaneously. Discovery of this interesting mode of regulation also raises the question as to whether or not miR-17–92 and miR-155 are specific miRNAs for TFH cell development and functions; however, this question remains to be clearly resolved. Moreover, several studies have demonstrated that miR-17–92 and miR-155 also regulate the Th1 responses against lymphocytic choriomeningitis virus infection and the generation of many other function T cell subsets (48495051), raising the general question as to how a single miRNA species is able to control so many different aspects of immune responses.
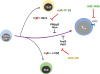 | Figure 1miRNA regulation of TFH cell differentiation. During the priming of naïve CD4+ T cells, miR-17–92, miR-155, and miR-146a are induced upon T cell receptor mediated stimulation. miR-17–92 and miR-155 positively regulate TFH cell differentiation via repression of negative regulators of TFH cells, Rora, Phlpp2, Pten, c-Maf, Peli1, and Fosl2. However, miR-146a tones down TFH cell functions by suppressing ICOS expression on the surface of effector TFH cells.
|
Recently, Lu et al. (52) elegantly explored this issue by generating Socs1 3′-untranslated region (UTR) mutant mice, which lack miR-155-mediated repression of only the Socs1 gene among the validated miR-155 targets. Previous studies demonstrated that miR-155 regulates Th17/Th2/Treg/TFH/natural killer (NK) cell generation and anti-viral immune responses using different experimental settings with miR-155 knockout mice (293132445053). These same studies indicated that Jarid2, Socs1, Fosl2, Peli1, Il23r, and c-Maf were functional targets, which are responsible for each of the observed phenotypes in the miR-155 knockout mice. Surprisingly, the Socs1 3′-UTR mutant mice phenocopied only a few phenotypes among those observed in the miR-155 knockout mice, such as NK cell expansion upon viral infection and enhanced Treg fitness in competitive settings (52). Considering all of these findings, Lu et al. (52) proposed that a single miRNA is capable of regulating several distinct immune responses by selecting a functional target in a cell type- and context-specific manner. This insightful conclusion can explain the variation observed within and between studies in 1 or 2 functional target genes even when using the same miR-155 knockout mice. However, this idea further brings forth a new and important question to resolve: how do miRNAs select the right target for the cell type and context?
Many research groups are currently pursuing answers to this abstruse question, including Xiao and colleagues (5455). Recently, this group executed an elegant systemic transcriptome and translatome analysis of miR-17–92-deficient or overexpressed primary B cells (56). Although the conclusions of this study were not able to provide clear answers to this question, they nevertheless provided important insights on this issue. First, there are more putative miR-17–92-binding sites in the pool of mRNAs than the total copy number of miR-17–92 species at the single-cell level, indicating that the presence of conserved miRNA-binding sites in the 3′-UTR of mRNA is not a sufficient condition to be repressed by any given miRNA. This implies the existence of certain mechanisms by which miRNAs select their targets. Second, the authors found that the validated target genes of miR-17–92 show distinct repression sensitivity to miR-17–92 in primary B cells. In other words, certain miR-17–92 target genes are repressed only after miR-17–92 are up-regulated in the B cells; however, those genes are not necessarily de-repressed when miR-17–92 are down-regulated. In contrast, another group of genes are responsive only when miR-17–92 are down-regulated; these 2 groups of genes rarely overlap with each other, indicating that functional target genes of miR-17–92 are probably selected on the basis of miRNA levels in B cells. Therefore, miRNAs might be able to select their appropriate target based on cell type and the context in accordance with the expression level of a given miRNA. Third, the secondary structure of mRNA was found to have a greater impact on its possibility of being selected as a target than other cis element criteria such as the length of the 5′-UTR, 3′-UTR, location, and numbers of miRNA-binding sites. Taken all together, these findings provide mechanistic insights into how the same miRNA species is able to choose different target genes in a cell type- and context-dependent manner.
The last few years has seen an explosion in research focused on defining the roles of miRNAs in the initiation and maintenance of the germinal center reaction. However, recent findings of the presence of a memory type of TFH cells implies the existence of another level of regulation of the germinal center reaction by memory TFH cells (57). Indeed, memory TFH cells rapidly respond to the second challenged pathogen and acquire TFH effector functions. This recall response of memory TFH cells is considered as a key event in long-lasting protective humoral immunity. Thus, revealing the mechanisms of the generation and maintenance of memory TFH cells might be crucial in the development of effective vaccines. Since CXCR5+ TFH-like cells are also characterized by plasticity with respect to their cell fate commitment upon stimulation, the roles of the memory type of TFH cells in various aspects of immunology are actively being explored (58). Therefore, defining the specific miRNA species and deciphering the molecular mechanisms of the miRNA-mediated regulation of memory TFH cell generation and functions will be of great interest, but remain to be elucidated.
 XML Download
XML Download