

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
B cell Ig diversity is achieved in 2 stages (1). The initial diversity is obtained by V(D)J rearrangement of the heavy chain and light chain, which involves Tdt-induced nucleotide addition and exonuclease-mediated deletion at the segment junctions (1). The second stage of diversification is achieved by somatic hypermutation (SHM) (1). SHM is initiated by activation-induced cytidine deaminase to form U/G mismatches followed by several possible fates, including C to T transition mutation, removal of the U-leading C→A/C/T mutation, or excision of neighboring bases and DNA synthesis (1). SHM preferentially targets C bases in the WRCY hot-spot motif (W=A or T, R=G or A, and Y=T or C), its reverse complementary sequence RGYW, or its variants, such as WRCH or DGYW. Since these hot-spot motifs are abundant within complementarity determining regions (CDRs) of Igs, SHM occurs more frequently in CDRs (1) and plays a critical role in the affinity maturation of B cell Igs.
Following the onset of recombinant antibody technology, artificial mutations were frequently introduced into CDRs to increase the affinity of an antibody (23). Development of experimentally-derived high affinity antibodies revealed that mutations of greater diversity can be introduced into CDRs to increase antibody affinity. Meanwhile, artificial mutation of the immunoglobulin framework region (FR) has been attempted for back-to-mouse mutations to restore affinity after grafting mouse CDRs into human Ig FR residues (4). However, if FR residues could be mutated without loss in affinity, improvements in physicochemical properties could be achieved more easily.
In this study, we investigated whether diverse mutations could be introduced into Ig FR residues while preserving antibody affinity. We selected a chicken antibody for this study because the chicken possesses only one functional VH and VL gene to make a functional antibody repertoire, even though shuffling with pseudogenes occurs (5). The FR residues of these pseudogenes have been reported to be homologous to the functional genes (5). Therefore, the chicken antibody is an ideal model system for comparing the FR of a natural repertoire to that of a synthetic one. This comparison is relatively less complicated to analyze in chicken than in mouse, rabbit, or human systems, all of which possess multiple VH and VL genes.
MATERIALS AND METHODS
Randomization of FR codons
A total of 161 PCRs were performed using degenerate primers to produce gene fragments encoding a portion of PC24 single chain Fv (scFv), an anti-prostate specific antigen (PSA) antibody (Genbank accession numbers KP764766 and KP764767), with an NNK degenerate codon (N=any nucleotide; K=G or T) at each of the 84 VH and 77 VL FR residues. The PCR products were fused to gene fragments encoding the remainder of the scFv gene with linker PCR to genes encoding a whole scFv. After restriction digestion with Sfi I, the genes were cloned into the pComb3XSS phagemid vector. The ligation products were used to transform competent ER2738 Escherichia coli (E. coli) to generate 161 scFv libraries as described previously (6). PC17 (another anti-PSA chicken antibody, unpublished) and VCM2 (an anti-VCAM-1 antibody, unpublished) were randomized at 16 framework residues to produce 16 scFv libraries in the same manner. All the amino acid of FR residues were numbered using Kabat numbering system (7).
Phage enzyme immunoassay
The scFv-displaying phages were rescued from titer plates after transformation and then used for phage enzyme immunoassay as described previously (6). Microtiter plates (Corning Inc., NY, USA) were coated with PSA (Fitzgerald Industries International, Acton, MA, USA) or recombinant human VCAM-1 (R&D Systems, Minneapolis, MN, USA) by adding 20 µl of protein in PBS (5 µg/ml) to each well followed by incubation overnight at 4°C. After blocking with 3% bovine serum albumin in PBS, the plate was then sequentially incubated with scFv-displaying phages in the culture supernatant, horseradish peroxidase (HRP)-conjugated mouse anti-M13 monoclonal antibody (GE Healthcare, Pittsburgh, PA, USA), and 2,2′-Azinobis [3-ethylbenzothiazoline-6-sulfonic acid]-diammonium salt (ABTS) substrate solution (Amresco, Solon, OH, USA) with intermittent washing using 0.05% Tween-20 in PBS. After incubating the plates at 37°C for 10 min, the optical density was measured at 405 nm using a microtiter plate reader (Labsystems, Barcelona, Spain).
Sequence analysis
Sequence analysis of phagemid clones was performed as described previously (6). Plasmid DNA was prepared using a small-scale plasmid preparation kit (Qiagen, Hilden, Germany). The pComb-F primer (ACA CTT TAT GCT TCC GGC TC) and pComb-R primer (CAA AAT CAC CGG AAC CAG AG) were used to determine the nucleotide sequence of VL and VH, respectively. The Accelrys Gene software program (Accelrys, San Diego, CA, USA) was used to analyze the sequence.
Generation of scFv-displaying phages with mutated H82, H82A, and H82B residues
PCR primers were designed to produce 70 mutants with different amino acids at these specific residues. PCR was performed to produce gene fragments encoding a portion of scFv, which was linked to the remainder of the scFv gene fragment by linker PCR as described above. The resulting scFv genes were digested with Sfi I and ligated into the pComb3X phagemid vector prior to transformation into ER2738 E. coli. The scFv-displaying phages were rescued from titer plates after transformation and used for phage enzyme immunoassay as described previously (6).
Enzyme immunoassay of chicken-human chimeric IgG1 with mutated H78 and L85 residues
Genes encoding 8 VH and 16 VL mutants were synthesized and cloned into an expression vector as described previously (6, 7). The IgG1 molecules were purified via protein A gel affinity chromatography following the instructions provided by the supplier (Repligen, Waltham, MA, USA). To check the affinity of the mutant IgG1s, enzyme immunoassay was performed using PSA-coated microtiter plates and HRP-conjugated anti-human IgG1 antibody (Thermo Fisher Scientific, Waltham, MA, USA) as described previously (6).
RESULTS
Versatility of the framework residues in chicken scFv
We constructed 161 sets of a scFv phage display library with randomly introduced amino acids at every FR residue within a chicken antibody (VH and VL consist of 84 and 77 residues, respectively) using degenerate oligomers. Each set contained a scFv randomized at a single FR residue. Subsequently, we randomly selected 92 clones from each library, rescued the scFv-displaying phage, and performed phage enzyme immunoassay. We defined positive clones as those exhibiting a 70% or greater signal over that of the original scFv phage rescued in parallel. Because in this setting, the absorbance value of positive clones and that of negative clones was different with statistically significance (p<0.05) among all sets of FR mutant library (Supplementary Fig. 1).
When the number of positive clones exceeded 20, only 20 clones were selected randomly for sequencing. Next, we classified the 20 amino acids into 5 groups based on their physicochemical properties as described previously (8): [G, A, P, V, L, I, M – nonpolar and aliphatic R groups], blue; [F, Y, W – aromatic R groups], purple; [S, T, C, N, Q – polar and uncharged R groups], green; [K, R, H – positively charged R groups], red; [D, E – negatively charged R groups], orange. Overall, 82 (50.9%), 48 (29.8%), 17 (10.6%), and 9 (5.6%) out of 161 residues could be replaced by a further 4, 3, 2, and 1 group of amino acid, respectively. Five FR residues (3.1%) of L23 (C), H36 (W), H86 (D), H104 (G), and H106 (G) were found to be replaced only with amino acids in the same group while maintaining its reactivity.
For comparison, we collected the amino acid sequences of 150 antigen-reactive chicken antibodies that we previously generated. Of these naturally-occurring chicken antibody FR, 1 (0.6%), 11 (6.8%), and 52 (32.3%) out of 161 FR residues were replaced by further 3, 2, and 1 group of amino acid, respectively. In case of 97 (60.3%) out of 161 residues, replacement with an amino acid from a different group was not found suggesting that naïve chicken antibody FR residues have limited sequence variations (Fig. 1).
 | Figure 1Comparing the interchangeability of 161 FR residues in PC24 scFv with the natural mutations found in naturally-occurring chicken antibodies. Amino acids were classified into 5 group of amino acid based on their physicochemical properties: [GAPVLIM], blue; [FYW], purple; [STCNQ], green; [KRH], red; [DE], orange. Positive rate was defined as the percentage of positive clones among 92 clones tested in phage enzyme immunoassay. The secondary structure of chicken antibody FR was adopted from the anti-phospho-tau antibody (PDB ID: 4GLR) (18). The sequences of 150 antigen-reactive chicken antibodies were collected, analyzed in the same manner, and presented as the natural mutation.
|
Effect of mutating 3 consecutive framework residues
Next, we investigated the effect of mutating 3 consecutive FR residues on the reactivity of a chicken antibody. We hypothesized that this arrangement of mutations would affect the antibody structure more significantly than mutation on 3 sporadic residues. Therefore, we mutated H82 residue, which is located at the end of the beta chain, along with H82A residue and H82B residue in the loop of the chicken anti-phospho-tau antibody (PDB ID: 4GLR), which is one of two chicken antibodies with a published 3-dimensional structure (9). Based on our previous data, the following substitutions could be made: H82 residue could be replaced with A, V, L, F, W, S, K, R, and H, which belong to 4 groups; H82A residue could be replaced with G, A, P, V, L, I, W, S, T, C, N, K, R, and E, which are members of 5 groups; and H82B residue could be substituted with G, P, V, L, I, F, W, S, T, C, N, Q, K, R, H, and D, which are also members of 5 groups.
A total of 70 mutants were created by replacing H82A residue with a different amino acid from the same group, substituting H82 residue with A, V, L, F, and W, and altering H82B residue to G, P, V, L, F, W, S, T, C, N, Q, K, R, and D. From these combinatorial mutants, we obtained 19 positive clones (Fig. 2). When both H82 residue and H82B residue were replaced with residues from different groups, only 6.3% (1 out of 16 mutants) were positive. Substitution of either H82 residue or H82B residue with an amino acid from a different group produced 19.4% (7 out of 36 mutants) positive clones. However, when both residues were mutated to amino acids from the same group, 61.1% (11 out of 18 mutants) were positive.
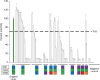 | Figure 2Effects of mutating 3 consecutive residues (H82, H82A, and H82B) on the reactivity of the scFv. With H82A FR residue were replaced into different amino acid in the same group of amino acid ([STCHQ], green), 70 different mutants were created by replacing H82 (A, V, L, F, and W) and H82B (G, P, V, L, F, W, S, T, C, N, Q, K, R, and D) FR residues simultaneously. Phage enzyme immunoassay was performed on the mutant scFv-displaying phage to determine its reactivity.
|
Versatility of the framework residues in 2 other chicken scFvs
To determine whether other chicken antibodies can tolerate mutations as effectively as the PC24 antibody, we tested 2 other antigen-reactive chicken antibodies. To accomplish this, we arbitrarily categorized the FR residues of PC24 into 2 classes: flexible and non-flexible. When a residue harbored 4 or 5 amino acid groups, we defined it as a flexible residue. Otherwise, the residues were classified as non-flexible residues. We selected 8 flexible residues (L5 [T], L6 [Q], L7 [P], L43 [A], H13 [T], H16 [G], H43 [K], and H82A [N]) and 8 non-flexible residues (L23 [C], L88 [C], H22 [C], H36 [W], H90 [Y], H92 [C], H103 [W], and H106 [G]). Subsequently, we performed mutational analysis on these residues in the two chicken antibodies. All 8 flexible residues were also found to be flexible in both chicken antibodies. Furthermore, 3 and 1 non-flexible residues were found to be flexible in the PC17 and VCM2 antibodies, respectively (Fig. 3). With these data, we concluded that the redundant nature of FR residues toward mutations is widely shared among different chicken antibodies.
 | Figure 3Versatility of 16 FR residues in 2 other chicken antibodies (PC17 and VCM2). Sixteen FR residues of 2 other antigen-reactive chicken antibodies, PC17 and VCM2 were randomized with the degenerate codon (NNK) generating 16 sets of scFv library. Amino acids were classified into 5 group of amino acid based on their physicochemical properties: [GAPVLIM], blue; [FYW], purple; [STCHQ], green; [KRH], red; [DE], orange. Positive rate was defined as the percentage of positive clones among 92 clones tested in each set of scFv library.
|
Versatility of the framework residues in chicken IgG1
Versatility chicken FRs are also monitored in IgG1 molecules. For this, H78 and L85 residues were selected and replaced with amino acids that provided positive signals in the scFv format (H78: G, A, L, M, F, S, Q, and H; L85: G, A, P, L, M, F, Y, W, S, T, N, Q, K, R, D, and E). All of the mutant IgG1 molecules bound to PSA in a dose-dependent manner (Fig. 4). These data demonstrated that the versatility of the chicken framework is not confined within the context of a scFv but could be extended to IgG1 molecules.

DISCUSSION
The extent to which SHM impairs FR structure and function has not been studied thoroughly. Previously, one article studied the effect of the random mutations on VH FR2 of a mouse monoclonal antibody (12). Only one out of 16 single residue mutants and 2 out of 8 double residue mutants had impaired antigen binding. When considering the fact that in the case of VH CDR2, 5 out of 20 single residue mutants and 6 out of 7 double residue mutants lost their reactivity, the possibility was proposed that VH FRs could accommodate many non-conservative changes. In this study, we showed that among 161 chicken antibody FR residues, 82 (50.9%), 48 (29.8%), 17 (10.6%), and 9 FR residues (5.6%) could be replaced by 4, 3, 2, and 1 group(s) of amino acids, individually, without significant loss of reactivity.
We evaluated the amino acid sequences of 150 naturally-occurring antigen-reactive chicken antibodies. Among these sequences, only 1 (0.6%), 11 (6.8%), and 52 (32.3%) FR residues were replaced by 3, 2, and 1 group(s) of amino acids, respectively. Furthermore, in a recent report analyzing the SHM of 1,269 unique naïve chicken VH FR residues, only 4 (4.9%), 3 (3.7%), 26 (32.1%), and 27 (33.3%) residues out of 81 residues were replaced by 4, 3, 2, and 1 groups, respectively (5). These results clearly demonstrate that the chicken antibody FR could tolerate much more mutational diversity than that observed in natural chicken antibodies.
One of the main reasons why the mutations in FR residues of natural chicken antibodies covers only a fraction of possible mutations is that the sequence composition of FRs is intrinsically less mutable than that of CDRs in human and mouse VH (11, 12). Moreover, the actual observed discrepancy in regional mutability in human VH is much larger; although the mechanism contributing to regional mutability is still not clear (12). Because the frequency of SHM in CDRs is limited to maintain the antigen reactivity in vivo, it is presumed that hypermutation in FRs would also be limited.
Another mechanism that could limit hypermutation of FRs is where mutations result in decreased stability and increased aggregation that would negatively affect secretion by B cells. The mutations in VH FR2 caused secretion defects in 24% (8 out of 33), a frequency twice that of VH CDR2 mutation (10). Interestingly, there are a few reports, in which stabilization of the variable region, especially in more unstable scFv or domain antibodies, could be achieved by introducing mutations in FRs. Substitution of hydrophobic FR residues with hydrophilic residues at surface-exposed positions increased the secretion of the scFv molecule in E. coli by reducing its aggregation potential (13). A single mutation in the VH FR2 (E43G) of a human scFv increased its stability (14). Furthermore, the stability of a human single domain VH antibody was dramatically increased by introducing 5 mutations on FRs (15). Because we tested the reactivity of the antibody in a phage-displayed format that should be less affected by the antibody stability, some unstable antibody clones could not be screened out.
Currently, we do not know if the versatility of FRs to accept mutations is a unique phenomenon of chicken antibody, which has only one functional VH and Vλ gene. However, our rather unexpected finding that chicken antibody FR residues can accommodate much more diverse sequence variations than those found in nature, including mutations of consecutive residues, is of significant benefit to the field of antibody engineering. Chicken FRs could be de-immunized with great versatility by introducing mutations (16). In addition, many FR residues could be substituted with cysteine or artificial amino acids to provide a point of conjugation with chemical entities including drugs, toxins, peptides, aptamers, isotopes or enzymes. In this case, the intrinsic characteristics of the antibody including in vivo pharmacokinetics, distribution, and effector functions would be less affected than antibodies conjugated through the Fc domain, which interacts with Fc receptors and complements.
 XML Download
XML Download