

 PDF
PDF Citation
Citation Print
Print



Abbreviations
CMI
cell-mediated immunity
FBS
fetal bovine serum
gE
glycoprotein E
HZ
herpes zoster
IE
immediate early
IEDB
Immune Epitope Data Base
IFN
interferon
MHC
major histocompatibility complex
mIL-33
murine IL-33
mIL-7
murine IL-7
OLP
overlapping peptide
PFU
plaque-forming unit
TNF
tumor necrosis factor
VV-gE
vaccinia virus expressing glycoprotein E
VZV
varicella-zoster virus
INTRODUCTION
Herpes zoster (HZ), or shingles, is caused by the reactivation of latent varicella-zoster virus (VZV) from the sensory ganglia (12). HZ develops when VZV-specific cell-mediated immunity (CMI) is decreased because of aging or immunosuppression (34). The estimated incidence of HZ is approximately 4 cases per 1,000 in those aged 50 to 60 years and increases to more than 10 cases per 1,000 for those aged >80 years (5). Lower VZV-specific CMI at HZ onset has been correlated with increased HZ severity and high incidence of complications (6). Therefore, VZV-specific CMI is thought to play a main role in controlling VZV reactivation and to contribute to protection against symptomatic HZ. Accordingly, the ability to elicit VZV-specific CMI is an important issue in the development of HZ vaccines (67).
A live attenuated VZV vaccine (Zostavax; Merck, Kenilworth, NJ, USA) has been available to prevent HZ since 2006. However, Zostavax has a limited vaccine efficacy depending on age (69.8% in those aged 50–59 years, 63.9% in those aged 60–69 years, 37.6% in those ≥70 years) (89). Moreover, the effectiveness of Zostavax decreases over time, from 68.7% in the first year after vaccination to approximately 4% by 8 years post-vaccination, which may account for the failure in eliciting long-term memory T-cell immunity after Zostavax vaccination (1011).
A recombinant subunit vaccine containing VZV glycoprotein E (gE) protein and the AS01B adjuvant system (Shingrix, HZ/su; GSK, Philadelphia, PA, USA) was recently introduced as an alternative and approved by the US Food and Drug Administration in October 2017. Two doses of HZ/su vaccine have clinically acceptable safety profiles and high vaccine efficacy regardless of age (96.6% in those aged 50–59 years, 97.4% in those aged 60–69 years, and 97.9% in those aged ≥70 years) (12). Studies have demonstrated that HZ/su elicits robust gE-specific CD4+ T cell and antibody responses, even in older adults and immunocompromised patients (71314). In a follow-up study, however, gE-specific CMI started to decline the first year after vaccination and fell to approximately 25% compared to the peak response between 36 and 72 months post-vaccination (15).
Since the first introduction in the early 1990s, DNA vaccines have been considered capable of inducing both cellular and humoral immune responses. In addition, DNA vaccines can easily be combined with other DNA plasmids encoding cytokines as molecular adjuvants (16). Notably, DNA vaccines have been suggested as an ideal therapeutic or prophylactic intervention, especially in chronic infectious diseases and cancers (17). DNA vaccines are currently being tested in clinical trials for the treatment of human papilloma virus-associated cervical cancer and breast cancer, and the prevention of human immunodeficiency virus and influenza infection (18192021).
IL-7 is a cytokine playing a significant role for T-cell homeostasis, particularly mediating the survival and expansion of naïve and memory T cells (2223). Previous reports demonstrated that in vivo administration of IL-7 results in the increase of T-cell numbers as well as antigen-specific, functional T-cell responses (242526). Furthermore, there are evidences that co-delivery of IL-7 as a vaccine adjuvant can enhance DNA vaccine-induced T-cell immune responses (2728). IL-33 is a member of the IL-1 cytokine family and acts as an endogenous ‘danger signal’ that triggers inflammation and promotes cell-mediated immune responses (29). Recently, IL-33 has been reported to enhance DNA vaccine-induced, anti-tumor or anti-viral T-cell immune responses (3031).
In the present study, we developed DNA vaccine candidates encoding VZV proteins and cytokine adjuvants, such as IL-7 and IL-33. We used gE, immediate early (IE) 63, and IE62 as vaccine antigens because they have been defined as immunodominant proteins (323334). We tested the immunogenicity in mice and found that HZ DNA vaccines induce VZV-specific functional T-cell responses, contributing to protective immunity in a surrogate challenge mouse model with vaccinia virus expressing gE (VV-gE). We further demonstrated that co-immunization with cytokine adjuvants enhanced HZ DNA vaccine-induced immunity. We observed that a single dose of HZ DNA vaccine had strong immune-boosting effects in mice with a history of previous infection by VV-gE. In summary, HZ DNA vaccines with cytokine adjuvants strongly elicit protective immunity.
MATERIALS AND METHODS
Animals
Five to six-week-old female C57BL/6 mice were purchased from Orientbio Inc. (Seongnam, Korea). All mice were maintained in specific pathogen-free conditions. Animal care and experimental procedures were performed with approval from the Animal Care Committee of Korea Advanced Institute of Science and Technology (KA2008-10).
DNA plasmids
Three different HZ DNA plasmids were generated using a modified pVAX1 mammalian expression vector and genes encoding gE (open reading frame, ORF 68), IE63 (ORF 63), and IE62 (ORF 62) of the pOka VZV strain. The genes encoding gE, IE63, and IE62 were genetically optimized for enhanced expression, including codon and RNA optimization, and a highly efficient immunoglobulin E leader sequence was added to facilitate expression. Each construct was synthesized commercially, sequence verified, and then the optimized gene was subcloned into a modified pVAX1 vector under control of the cytomegalovirus immediate-early promoter (GeneArt, Regensburg, Germany). The 3 HZ DNA plasmids were designated pVAX1-gE, pVAX1-IE63, and pVAX1-IE62, respectively. For cytokine DNA plasmids, the gene sequences for murine IL-7 (mIL-7) and murine IL-33 (mIL-33) were obtained from the NCBI GenBank Database and the genes synthesized. They were subcloned into a modified pVAX1 vector, designated pVAX1-mIL-7 and pVAX1-mIL-33.
Immunization
Five to 6-week-old female C57BL/6 mice were intramuscularly immunized 3 times at 2-week intervals with 30 µg pVAX1-gE, pVAX1-IE63, or pVAX1-IE62 in 100 µl of PBS, followed by in vivo electroporation using a CELLECTRA device (Inovio Pharmaceuticals, Inc., Plymouth Meeting, PA, USA) on the external thigh. Control mice were immunized with 30 µg modified pVAX1 plasmid (pVAX1). Cytokine adjuvants (30 µg pVAX1-mIL-7 or pVAX1-mIL-33) were used simultaneously with HZ DNA vectors. Two weeks after the last immunization, mice were sacrificed, and their spleens obtained.
For the experiment of mice with a history of VV-gE infection, 6-week-old mice were intraperitoneally infected with 1×107 plaque-forming units (PFUs) of VV-gE in 100 µl PBS, maintained for 2 months, and DNA vaccination performed. Mice were sacrificed 2 weeks after the immunization and the spleens obtained.
Overlapping peptides
Overlapping peptides (OLPs; Mimotopes Pty Ltd., Melbourne, Australia) were synthesized as 15-mers overlapping by 10 amino acids to cover the whole amino acid sequence of gE, IE63, and IE62. Lyophilized peptides were solubilized in 5% DMSO (Sigma-Aldrich, St. Louis, MO, USA). OLPs were pooled as follows: gE-11-41, gE-242-82, gE-383-123, IE63-11-27, IE63-228-54, IE62-11-37, IE62-238-74, IE62-375-111, IE62-4112-148, IE62-5149-185, IE62-6186-223, and IE62-7224-261. The concentration of each peptide in the pools was 25 µg/ml, and finally diluted to 1 µg/ml in the splenocyte culture.
Interferon (IFN)-γ ELISPOT assays
IFN-γ ELISpot assays were performed as described previously to measure antigen-specific IFN-γ secretion (35). The spleens of immunized mice were collected in RPMI-1640 medium (WelGENE, Daegu, Korea) supplemented with 5% fetal bovine serum (FBS) and 1x penicillin-streptomycin, mechanically mashed, and filtered using 40 µm strainers. After centrifugation, cells were treated with RBC lysis buffer (BioLegend, San Diego, CA, USA) for 5 min at room temperature, washed, and then resuspended (0.5×106/well) in RPMI medium. Splenocytes were stimulated with OLP pools for 24 h. PMA (10 ng/ml, Sigma-Aldrich) and ionomycin (500 ng/ml, Sigma-Aldrich) were used as a positive control and 5% DMSO (Sigma-Aldrich) as a negative control. In addition to OLP pools, VZV-infected cell lysate (Microbix Biosystems, Mississauga, ON, Canada) was used as a stimulant, and non-infected MRC5 lysate (Microbix Biosystems) was used as a negative control. Spots were enumerated using an automated ELISpot reader (AID GmbH, Strassberg, Germany) and the number of specific spots calculated by subtracting the spot number in negative control wells from the spot number in OLP pool- or VZV lysate-stimulated wells.
Intracellular cytokine staining
Splenocytes were ex vivo stimulated with the immunodominant OLP pool, gE-11-41, IE63-228-54, and IE62-375-111 of gE, IE63, or IE62, respectively. To measure the degranulation activity of T cells, anti-CD107a-BV421 (1D4B; BioLegend) was added during the stimulation. Brefeldin A (GolgiPlug, BD Biosciences, San Jose, CA, USA) and monensin (GolgiStop, BD Biosciences) were added to the culture 1 h after the stimulation, and the culture was further maintained for 5 h. The Live/Dead Fixable Red Dead Cell stain kit (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA), anti-CD3-Alexa Fluor 700 (500A2; eBioscience, Thermo Fisher Scientific), anti-CD4-BV786 (GK1.5; BD Biosciences), and anti-CD8-BV510 (53-6.7; BD Biosciences) were used for immunostaining at 4°C for 20 min. For intracellular staining, cells were permeabilized and fixed using the BD Cytofix/Cytoperm kit (BD Biosciences) at 4°C for 20 min, followed by staining with anti-IFN-γ-APC (XMG1.2; BioLegend), anti-tumor necrosis factor (TNF)-FITC (MP6-XT22; eBioscience), and anti-IL-2-PE (JES6-5H4; eBioscience) at 4°C for 20 min. All data were collected using a LSRΙΙ flow cytometer (BD Biosciences) and analyzed using FlowJo software (Treestar, Ashland, OR, USA). Boolean gating was performed using FlowJo software to analyze the polyfunctionality of T cells, and charts were visualized by SPICE software (https://niaid.github.io/spice/).
Epitope mapping and major histocompatibility complex (MHC) class Ι dextramer staining
Because pVAX1-IE62-immunized mice exhibited a strong T-cell response against the IE62-375-111 OLP pool, we mapped T-cell epitopes within IE62-375-111. We performed IFN-γ ELISpot assays with splenocytes from pVAX1-IE62-immunized mice by stimulating them with a single peptide (15-mer; total 37 peptides). Peptides with high numbers of IFN-γ spots were identified, and the epitope sequence of high-affinity binding to mouse MHC class Ι, H-2Db was identified using Immune Epitope Data Base (IEDB) website. The epitope peptide, RALINLIYC, was used for synthesis of H-2Db dextramer (H-2Db IE62527-535 dextramer, Immunedex, Copenhagen, Denmark). Two weeks after the last immunization of pVAX1-IE62 with pVAX1-mIL-7 or pVAX1-mIL-33, splenocytes were stained with PE-conjugated H-2Db IE62527-535 dextramer for 10 min at room temperature in the presence of anti-mouse CD16/32 (2.4G2; BD Biosciences), followed by washing with FACS buffer (PBS containing 1% FBS and 0.05% sodium azide). For live/dead and surface staining, the following reagent and antibodies were used: Live/Dead Fixable Red Dead Cell stain kit (Invitrogen), anti-CD19-PE-CF594 (1D3), anti-CD4-APC-Cy7 (GK1.5), anti-CD8-BV510 (53-6.7), anti-KLRG1-Alexa Fluor 488 (2F1), anti-CD62L-BV605 (MEL-14; all from BD Biosciences), anti-CD44-BV650 (IM7), anti-CD69-BV711 (H1.2F3), anti-CD43-APC (1B11), anti-CD127-BV421 (A7R34; all from BioLegend), anti-CD27-PE-Cy7 (LG.7F9), and anti-CD3-Alexa Fluor 700 (500A2; all from eBioscience).
Recombinant vaccinia virus challenge and plaque assays
Recombinant VV-gE was used for in vivo challenge of a surrogate virus. Immunized mice were challenged intraperitoneally with 1×107 PFU VV-gE in 100 µl PBS 14 days after the last vaccination. Mice were sacrificed 2 and 5 days after the challenge. Ovaries were harvested, placed in DMEM medium (WelGENE), and homogenized using Precellys 24 homogenizer (Bertin, Montigny-le-Bretonneux, France). Supernatants were collected and frozen at −70°C until plaque assays were performed. 143TK cells were grown on rat tail type Ι collagen (56 µg/well; Corning Inc., Corning, NY, USA)-coated 6-well plates to confluence. Ten-fold dilutions of the supernatants were added to the monolayer of 143TK cells and incubated at 37°C for 1 h. After removing diluted supernatants, DMEM supplemented with 5% FBS and 1× penicillin-streptomycin was added and left overnight. Staining was performed with 0.1% crystal violet after fixation with 1% paraformaldehyde. Plaques were counted and total viral load per ovary calculated.
Statistical analysis
Statistical analyses were performed using Prism software 6.0 (GraphPad Software Inc., La Jolla, CA, USA). Unpaired Student t-tests (Mann-Whitney U test) were used to compare the results between two groups. To analyze the significance of polyfunctionality, 2-way ANOVA multiple comparisons were performed. Differences were considered significant when p<0.05 with a 95% confidence level.
RESULTS
HZ DNA vaccines induce VZV protein-specific T-cell immune responses
To investigate the immunogenicity of HZ DNA vaccines, C57BL/6 (B6) mice were intramuscularly immunized with pVAX1-gE, pVAX1-IE63, or pVAX1-IE62, followed by in vivo electroporation according to the schedule presented in Fig. 1A. When splenocytes from the immunized mice were stimulated with OLP pools corresponding to the immunized VZV ORF, IFN-γ ELISpot assays showed that all three DNA vaccines elicited robust T-cell responses against the antigen protein (Fig. 1B). Strong T-cell responses were also observed when splenocytes were stimulated with VZV-infected cell lysate (Fig. 1B). Next, we identified which T-cell subset responds to VZV proteins after HZ DNA vaccination by performing intracellular cytokine staining for IFN-γ, TNF, and IL-2. In the assays, splenocytes were stimulated with the immunodominant OLP pool for each VZV antigen protein as follows: gE-11-41, IE63-228-54, and IE62-375-111. pVAX1-gE elicited gE-specific production of IFN-γ, TNF, and IL-2 from CD4+ T cells, but not from CD8+ T cells (Fig. 1C). Similarly, pVAX1-IE63 induced a CD4+ T-cell response, but not CD8+ T-cell response, although IL-2 was not produced from CD4+ or CD8+ T cells (Fig. 1C). In contrast, pVAX1-IE62 elicited IE62-specific production of IFN-γ and TNF from both CD4+ and CD8+ T cells, and a high frequency of CD8+ T cells produced IFN-γ and TNF in response to IE62 (Fig. 1C). Thus, HZ DNA vaccines induced robust T-cell responses mediated mainly by either CD4+ or CD8+ T cells depending on the antigen protein.
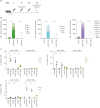 | Figure 1T-cell immunogenicity of HZ DNA vaccines in C57BL/6 mice. Five to six-week-old female C57BL/6 mice were intramuscularly immunized three times at 2-week intervals with 30 µg pVAX1-gE, pVAX1-IE63, or pVAX1-IE62 in 100 µl of PBS, followed by in vivo electroporation (n=5 for each group). The control mice were immunized with 30 µg pVAX1 (n=3). The mice were sacrificed 2 weeks after the last immunization. (A) Immunization schedule. (B) IFN-γ ELISpot assays were performed to detect antigen-specific, IFN-γ-producing T cells by stimulating splenocytes from immunized mice with OLP pools corresponding to the immunized VZV ORF or VZV-infected cell lysate. (C) Intracellular cytokine staining for IFN-γ, TNF, and IL-2 was performed to detect antigen-specific, cytokine-producing CD4+ and CD8+ T cells by stimulating splenocytes from immunized mice with the immunodominant OLP pool for each VZV antigen protein (gE-11-41, IE63-228-54, and IE62-375-111). Data are presented as means and standard deviations.
*p<0.05.
|
IL-7 and IL-33 adjuvants enhance HZ DNA vaccine-induced T-cell immunity
Studies have shown that co-delivery of plasmids encoding cytokines can augment immune responses in DNA vaccination (36). In the present study, we tested IL-7 and IL-33 as molecular adjuvants in HZ DNA vaccination. Co-administration of plasmids encoding IL-7 or IL-33 have been reported to enhance DNA vaccine-induced T-cell responses (27283031). Mice were immunized with pVAX1-gE combined with pVAX1-mIL-7 or pVAX1-mIL-33, and gE-specific T-cell responses evaluated by intracellular cytokine staining. As a result, pVAX1-gE immunization with both pVAX1-mIL-7 and pVAX1-mIL-33 elicited the strongest T-cell responses (Fig. 2A and B). Compared to pVAX1-gE alone, pVAX1-gE with both pVAX1-mIL-7 and pVAX1-mIL-33 significantly increased the TNF or IL-2 production by CD4+ T cells (Fig. 2B). pVAX1-gE with both pVAX1-mIL-7 and pVAX1-mIL-33 also significantly increased IFN-γ, TNF, or IL-2 production by CD4+ T cells compared to pVAX1-gE with pVAX1-mIL-7 (Fig. 2B). In polyfunctional T-cell analysis for IFN-γ, TNF, and IL-2, pVAX1-gE with both pVAX1-mIL-7 and pVAX1-mIL-33 tended to increase the percentage of triple- or double-positive T cells (Fig. 2C and D). In particular, pVAX1-gE with both pVAX1-mIL-7 and pVAX1-mIL-33 significantly enhanced the frequency of IFN-γ+TNF+CD4+ T cells compared to pVAX1-gE alone or pVAX1-gE with plasmids for a single cytokine (Fig. 2D).
 | Figure 2Adjuvant effects of IL-7 and IL-33 on HZ DNA vaccine-induced CD4+ T-cell immunity. Five to 6-week-old female C57BL/6 mice were intramuscularly immunized three times at 2-week intervals with 30 µg pVAX1-gE combined with 30 µg pVAX1-mIL-7 or pVAX1-mIL-33 in 100 µl of PBS, followed by in vivo electroporation (n=5 for each group). The control mice were immunized with 30 µg pVAX1 (n=3). Intracellular cytokine staining for IFN-γ, TNF, and IL-2 was performed to detect gE-specific, cytokine-producing CD4+ T cells by stimulating splenocytes from immunized mice with the gE-11-41 OLP pool. (A) Representative FACS dot plots. (B) Percentages of each cytokine-producing cell among CD4+ T cells for IFN-γ, TNF, and IL-2. Data are presented as means and standard deviations. (C) Pie graphs show the fraction of CD4+ T cells positive for a given number of functions. (D) CD4+ T cell polyfunctionality analyzed by every possible combination of functions and compared among groups. Means are presented.
*p<0.05; ***p<0.001.
|
We also tested IL-7 and IL-33 as molecular adjuvants in the pVAX1-IE62-induced CD8+ T-cell response. First, we attempted to identify a CD8+ T-cell epitope peptide in the immunodominant IE62 OLP pool, IE62-375-111, from pVAX1-IE62-immunized mice by performing IFN-γ ELISpot assays with single OLP stimulation. This assay revealed two subsequent OLPs, DPARQYRALINLIYC and YRALINLIYCPDRDP, as strong stimulants (Fig. 3A). In the overlapped sequence, we picked up RALINLIYC (IE62527-535) as a high-affinity binder to H-2Db using the IEDB website, and finally synthesized H-2Db IE62527-535 dextramer. Using this dextramer, we successfully detected IE62527-535-specific CD8+ T cells in pVAX1-IE62-immunized mice (Fig. 3B). Mice were then immunized with pVAX1-IE62 combined with pVAX1-mIL-7 or pVAX1-mIL-33, and the frequency of IE62527-535-specific CD8+ T cells was evaluated by dextramer staining. As a result, pVAX1-IE62 immunization with both pVAX1-mIL-7 and pVAX1-mIL-33 elicited significantly higher CD8+ T-cell responses than pVAX1-IE62 alone (Fig. 3C). Taken together, the results indicate that co-administration of both pVAX1-mIL-7 and pVAX1-mIL-33 significantly enhanced both CD4+ and CD8+ T-cell responses in HZ DNA vaccination.
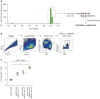 | Figure 3Adjuvant effects of IL-7 and IL-33 on HZ DNA vaccine-induced CD8+ T-cell immunity. (A, B) Five to six-week-old female C57BL/6 mice were intramuscularly immunized three times at 2-week intervals with 30 µg pVAX1-IE62 in 100 µl of PBS, followed by in vivo electroporation. IFN-γ ELISpot assays were performed by stimulating splenocytes from immunized mice with a single OLP (OLP #75–#111) from the immunodominant IE62-375-111 pool, and OLPs with high numbers of IFN-γ spots were identified. The epitope sequence, RALINLIYC (IE62527-535), with high-affinity binding to mouse MHC class Ι, H-2Db was identified by IEDB analysis, and the corresponding H-2Db IE62527-535 dextramer synthesized (A). IE62527-535-specific CD8+ T cells were detected by staining splenocytes from the pVAX1-IE62-immunized mice using H-2Db IE62527-535 dextramer (B). (C) Five to six-week-old female C57BL/6 mice were intramuscularly immunized three times at 2-week intervals with 30 µg pVAX1-IE62 combined with 30 µg pVAX1-mIL-7 or pVAX1-mIL-33 in 100 µl of PBS, followed by in vivo electroporation (n=3 for each group). Percentages of IE62527-535-specific cells among spleen CD8+ T cells were analyzed by staining splenocytes using H-2Db IE62527-535 dextramer and compared among groups. Data are presented as means and standard deviations.
*p<0.05.
|
T-cell responses elicited by HZ DNA vaccination are protective against a surrogate infection
There is no appropriate mouse model for HZ because of host restriction of VZV. To evaluate whether HZ DNA vaccination induced protective immunity, we used VV-gE as a surrogate virus in challenge experiments. First, mice were immunized with pVAX1-gE with pVAX1-mIL-7 or pVAX1-mIL-33. Next, the mice were challenged with 1×107 PFU VV-gE according to the schedule in Fig. 4A. The PFUs of VV-gE in the ovaries was determined 2 and 5 days after the challenge. Mice immunized with pVAX1-gE exhibited significant viral reduction compared to mice immunized with pVAX1 control plasmids, regardless of co-administration with pVAX1-mIL-7 or pVAX1-mIL-33 (Fig. 4B). Compared to pVAX1-gE alone, pVAX1-gE with pVAX1-mIL-33, or both pVAX1-mIL-7 and pVAX1-mIL-33 significantly decreased the viral titer 2 days post-infection (Fig. 4B). These data indicate that HZ DNA vaccination successfully induces protective immunity with additive benefits of IL-7 and IL-33 adjuvants.
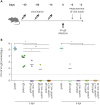 | Figure 4HZ DNA vaccine-induced protective immunity against a surrogate infection. Five to 6-week-old female C57BL/6 mice were intramuscularly immunized three times at 2-week intervals with 30 µg pVAX1-gE combined with 30 µg pVAX1-mIL-7 or pVAX1-mIL-33 in 100 µl of PBS, followed by in vivo electroporation (n=5 for each group). The control mice were immunized with 30 µg pVAX1 (n=3). The immunized mice were intraperitoneally challenged with 1x107 PFU VV-gE 2 weeks after the last immunization. (A) The schedule of immunization and viral challenge is presented. (B) Viral loads (PFU) in ovaries were measured 2 and 5 days post-infection. Means are presented.
*p<0.05; **p<0.01.
|
Single dose of HZ DNA vaccines boosts T-cell immunity in mice previously exposed to VV-gE
Zoster vaccines are administered to hosts who are latently infected with VZV and already have VZV-specific T-cell immunity, though it was attenuated by aging or immunosuppression. To mimic this situation, mice were infected with 1×107 PFU VV-gE. After recovery from the VV-gE infection, the mice were immunized with a single dose of HZ DNA vaccine according to the schedule in Fig. 5A. When T-cell responses were evaluated by IFN-γ ELISpot assays with stimulation of gE OLP pools, VV-gE infection did not result in detectable levels of T-cell responses 2 months after the infection. However, a single dose of pVAX1-gE strongly boosted gE-specific T-cell immunity regardless of the co-administration with pVAX1-mIL-7 or pVAX1-mIL-33 (Fig. 5B). Similar results were observed when splenocytes were stimulated by VZV-infected cell lysate (Fig. 5B). We also performed intracellular cytokine staining in the same setting. A single dose of pVAX1-gE immunization robustly boosted gE-specific production of IFN-γ, TNF, and IL-2 by CD4+ T cells regardless of the co-administration with pVAX1-mIL-7 or pVAX1-mIL-33, although VV-gE infection without HZ DNA immunization did not result in detectable levels of cytokine production by CD4+ T cells (Fig. 5C). These data indicate that even a single dose of HZ DNA vaccine robustly boosts VZV-specific T-cell immune responses in mice previously infected with VV-gE.
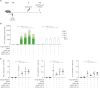 | Figure 5Immune-boosting effects of HZ DNA vaccines in mice previously infected with VV-gE. Five to 6-week-old female C57BL/6 mice were intraperitoneally infected with 1x107 PFU VV-gE. Sixty days later, the mice were intramuscularly immunized with 30 µg pVAX1-gE combined with 30 µg pVAX1-mIL-7 or pVAX1-mIL-33 in 100 µl of PBS, followed by in vivo electroporation (n=5 for each group). The control mice were immunized with 30 µg pVAX1 (n=5). The mice were sacrificed 2 weeks after the immunization. (A) Schedule of infection and immunization. (B) IFN-γ ELISpot assays were performed to detect antigen-specific, IFN-γ-producing T cells by stimulating splenocytes from the immunized mice with gE OLP pools (gE-11-41, gE-242-82, and gE-383-123) or VZV-infected cell lysate. (C) Intracellular cytokine staining for IFN-γ, TNF, and IL-2 was performed to detect antigen-specific, cytokine-producing CD4+ and CD8+ T cells by stimulating splenocytes from immunized mice with the gE-11-41 OLP pool. Data are presented as means and standard deviations.
**p<0.01.
|
DISCUSSION
Here, we demonstrated that HZ DNA vaccination with IL-7 and IL-33 molecular adjuvants elicited protective T-cell immunity. Moreover, we showed that a single dose of HZ DNA vaccination robustly boosted VZV-specific T-cell responses in mice with a history of previous infection with VV-gE. Zostavax is a live attenuated vaccine that has been used in the elderly. However, the vaccine has limitations, including relatively low rates of prevention and declining vaccine efficacy with age. In addition, its use is contraindicated in immunocompromised people (37). HZ/su is a recombinant subunit vaccine containing gE protein and the AS01B adjuvant. HZ/su exhibits long-term, high vaccine efficacy regardless of the age of the host. However, Hu/su significantly induces gE-specific T-cell responses only when two-dose vaccination is performed (7). Therefore, HZ vaccines that vigorously induce VZV-specific T-cell responses with a single-dose vaccination and can be used safely for immunocompromised patients need to be developed.
We evaluated the T-cell immunogenicity of HZ DNA vaccines encoding VZV gE, IE63, or IE62 and found that HZ DNA vaccines robustly induce VZV-specific T-cell responses. Furthermore, the T-cell immunogenicity of HZ DNA vaccines was enhanced by co-administration of DNA plasmids encoding IL-7 and IL-33. Previous studies have shown that co-delivery of plasmids encoding various cytokines substantially enhances DNA vaccine-induced immune responses in diverse settings (203839). In particular, plasmids encoding IL-7 or IL-33 have been shown to significantly enhance DNA vaccine-induced T-cell responses (27283031). In the current study, we evaluated the adjuvant effects of the combination of IL-7 and IL-33 for the first time and found that combined administration of IL-7 and IL-33 plasmids strongly enhanced HZ DNA vaccine-induced T-cell responses (Fig. 2). The strongest effect of the combination of IL-7 and IL-33 was also observed in the VV-gE challenge experiment, particularly at the early phase of infection (2 days post-infection; Fig. 4), indicating that HZ DNA vaccines co-administered with both IL-7 and IL-33 plasmids contribute to immediate protective immunity. This is important because high levels of immediate T-cell responses at the onset of HZ could attenuate the severity of HZ and the incidence of complications (6).
In the current study, we tried to assess the immune-boosting effects of HZ DNA vaccination in mice with a history of previous infection with VV-gE to mimic clinical situations, in which most people are latently infected with VZV and have pre-existing VZV-specific immunity (40). Similar experiments were performed during the development of HZ/su. Mice with a history of subcutaneous administration of live attenuated VZV were immunized with HZ/su; VZV-specific T-cell responses were strongly induced by vaccination with 2 doses of HZ/su (41). In contrast, a single dose of HZ DNA vaccine significantly increased VZV-specific T-cell responses in mice previously infected with VV-gE (Fig. 5). This result suggests that a single-dose regimen may be enough to prevent the development of HZ when HZ DNA vaccines are clinically developed.
The goal of HZ vaccination is to increase VZV-specific T-cell responses in order to prevent HZ and its complications. Here, we showed that HZ DNA vaccines induce strong VZV-specific T-cell responses, which are further enhanced by co-administration of plasmids for molecular cytokine adjuvants, such as IL-7 and IL-33. We also showed that HZ DNA vaccine-induced immunity is protective against virus infection using a surrogate virus. Importantly, a single dose of HZ DNA vaccination robustly boosts T-cell responses in mice with a history of previous infection by a surrogate virus. Thus, HZ DNA vaccination could be a promising strategy for prevention of HZ.
 XML Download
XML Download