

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The immune system consists of an intricate network of organs, tissues, cells, and molecules responsible for maintaining the body's homeostasis and responding to the host's aggression state. Innate immunity operates as the organism's first line of defense in conjunction with adaptive immunity and is characterized by a rapid response to pathogen invasion. The main cellular components of the innate immune system include phagocytic leukocytes, dendritic cells, and a special type of lymphocytes called natural killer (NK) cells (1). NK cells have been identified in both lymphoid and non-lymphoid tissues and can quickly reach target organs under pathologic conditions. NK cells are important in a host's immune response in the early stage of cancer development, metastasis, and/or virus infection (23). NK cells are principally in a resting phase but infiltrate tissues in cases of activation (4). Several soluble mediators, including cytokines, chemokines, and soluble receptor ligands, as well as cell-to-cell interactions, are involved in the regulation of NK cell functions (5). Among the cytokines, type I interferon (IFN), IL-12, and IL-18 are potent activators of NK cell effector function (6). Additionally, IL-2 promotes NK cell proliferation, cytotoxicity, and, to some extent, cytokine secretion (7). Upon activation with various stimuli, including the engagement of surface receptors such as natural cytotoxicity receptors and NKG2D, NK cells release cytotoxic granules and secrete cytokines such as IFN-γ that induce target cell death (8).
Since the 1970s, many bioactive peptides derived from food protein hydrolysates have been studied as potential nutraceuticals (91011). Dietary proteins and amino acids are important for immune function, some of which act as modulators of the immune response (1213). Among the dietary proteins, silk peptide has been proposed as a bioactive supplement (14). Silk peptide has been ingested as a healthy functional food in Asian countries; they are natural biomolecules used in powder and extract forms in diverse pharmacological capacities as well as in biomedical and biotechnological fields (15). Silk peptide biopolymers from the cocoons produced by silkworms for protection from the environment during metamorphosis to the mature moth stage (16). The silk peptide component is similar to proteins such as collagen, elastin, keratin, fibroin, and sporgin and is an important ingredient of cocoons (17). Silk peptide displays diverse bioactivities, such as anti-inflammatory, immune-regulatory, anti-tumor, anti-viral, and anti-bacterial (181920). Importantly, silk fibroin peptides and the silk protein sericin inhibit the proliferation of human lung cancer cells, induce apoptosis, inhibit the formation of colorectal tumors, and exert anti-cancer effects against cancer cells (212223).
Earlier investigations demonstrated that silk peptide activates macrophages, enhance their phagocytic ability, and stimulates the production of pro-inflammatory cytokines (24). Silk peptide also induces strong tumor necrosis factor production and weaker production of IL-1β and IL-6 in RAW 264.7 cells (25). However, there is currently no available information on the effects of silk peptide on NK cell activity. Given that silk peptide exerts anti-tumor activity, we investigated the function of silk peptide on the activation of NK cells, which remove abnormal cells including tumor cells. We confirmed that silk peptide enhances NK cell activity in vitro and ex vivo. In addition, oral administration of silk peptide significantly enhanced the frequency and target cell cytolytic activity of NK cells.
MATERIALS AND METHODS
Animal experiments
Six-week-old female C57BL/6 mice were purchased from the Koatech Laboratory Animal Center (Pyeongtaek, Korea) and housed under specific pathogen-free conditions. Mice were divided into five experimental groups with five mice per group. Mice were freely fed feeds containing silk peptide (2 g feed/day/20 g mouse) for 2 months, and each group of mice was administered 0, 750, 1,500, 3,000, and 7,500 mg silk peptide/kg body weight/day. Animal experiments were approved by the Institutional Animal Care and Use Committee of Chonbuk National University (approval No. CBNU 2018-074) and followed the guidelines suggested by the committee.
Preparation of acid-hydrolyzed silk peptide
The silk peptide used in this study was prepared from cocoons of Bombyx mori L. and is an ingredient for manufacturing commercial products of Worldway Co., Ltd. (Sejong, Korea). Briefly, silk worm cocoons were hydrolyzed, followed by neutralization, decolorization, filtration, desalting, concentration, and drying to obtain a pale yellow powder. Silk peptide (5 mg) was dissolved in 20 mM HCl solution (100 mL) and derivatized. Derivatized samples were quantitated on a Waters 2695 Alliance HPLC System (Waters Co., Milford, MA, USA) and a Waters 2475 Multi-λ Fluorescence Detector. The amino acid composition of the silk peptide used in this study was alanine (20.0%), glycine (31.3%), serine (15.3%), and others (17.95%).
Experimental materials
Unless otherwise specified, the chemicals and laboratory wares used in this study were purchased from Sigma Chemical Co. (St. Louis, MO, USA) and SPL Life Sciences (Pocheon, Korea), respectively. Fetal bovine serum and RPMI-1640 medium were obtained from Hyclone Laboratories (Logan, UT, USA). The mouse lymphoma YAC-1 cell line was kindly provided by Kim C (Inha University, Incheon, Korea). The Cytometric Bead Array Mouse T helper (Th)1/Th2 Cytokine Kit for cytokine detection was purchased from BD Pharmingen (Franklin Lakes, NJ, USA). 3H-TdR was purchased from Perkin-Elmer Life and Analytical Sciences (Shelton, CT, USA).
Abs and flow cytometric analyses
The following Abs against mouse molecules were purchased from Miltenyi Biotec Inc. (Bergisch Gladbach, Germany): CD3-FITC, NK1.1-PerCP-Vio700, CD11b-allophycocyanin (APC)-Vio770, CD27-PE-Vio770, CD107a-APC, IFN-γ-PE, CD4-PerCP-Vio700, and CD8a-APC. 7-Amino-actinomycin D (7-ADD) and FITC-conjugated annexin V were purchased from BD Biosciences (San Jose, CA, USA). Flow cytometric analysis was performed using a Cytoflex flow cytometer (Beckman Coulter, Inc., Brea, CA, USA) and data analysis was performed with CytExpert software (Beckman Coulter, Inc.).
Lymphocyte proliferation
Cell proliferation was determined using the 3H-TdR incorporation assay. Briefly, 1×105 cells were seeded into each well of a 96-well plate together with 100 μl of concanavalin A (Con A; 5 μg/ml), lipopolysaccharide (LPS; 2 μg/ml), or IL-2 (50 ng/ml). Following incubation at 37°C for 48 h in a CO2 incubator, 0.5 μCi of 3H-TdR was added to each well. After a 12-h chase incubation, the cells were collected with a 96-well cell harvester (Inotech, Dottikon, Switzerland), and the incorporated tritium content was counted using a 1450 Microbeta liquid scintillation counter (Perkin-Elmer Life and Analytical Sciences). Stimulation indices were calculated by dividing the tritium incorporation (cpm) in cells treated with the sample with the incorporation in control cells treated with phosphate-buffered saline.
Analysis of cytokine expression
To analyze cytokine production in splenocytes, cells (2×105) were distributed into each well of a 24-well plate and stimulated with phorbol 12-myristate 13-acetate (PMA; 50 ng/ml) and ionomycin (0.5 μg/ml) for 4 h. The culture medium was collected after stimulation and the expression levels of various cytokines were measured using a CBA Mouse Th1/Th2 Cytokine Kit (BD Pharmingen) according to the manufacturer's recommendations. The results were analyzed using FCAP Array software (BD Biosciences). Cytokine concentrations were calculated using a standard curve generated from cytokine standards.
Determination of NK cell activity
NK cells were purified from splenocytes prepared from three mice via magnetic beads using a NK Cell Isolation Kit II (Miltenyi Biotec Inc.). The degree of NK cell maturation was determined through flow cytometry by measuring the expression levels of CD3, NK1.1, CD27, and CD11b. To measure NK cell activation, the cell surface expression of CD107a, a marker for NK cell degranulation, and intracellular IFN-γ expression were determined (2627). Briefly, NK cells were stimulated with YAC-1 target cells at a ratio of 5:1 or without YAC-1 target cells in the presence of brefeldin A (10 μg/ml) and monensin (6 μg/ml) and incubated at 37°C for 6 h in a CO2 incubator. To measure the level of NK cell degranulation, FITC-conjugated anti-CD107a Ab was added and the level was measured in CD3−NK1.1+ cells. In addition, intracellular IFN-γ production was determined in CD3−NK1.1+ cells via flow cytometry.
To measure NK cell-mediated cytotoxicity, YAC-1 tumor target cell lysis was measured as described previously (28). Briefly, YAC-1 target cells were labeled with Paul Karl Horan (PKH)-26 and mixed with effector cells at various effector to target ratios. Cells were plated and incubated for 4 h at 37°C in a CO2 incubator. Following incubation, 7-ADD and FITC-annexin V were added to determine the level of apoptosis induced in target cells via flow cytometry.
Statistical analyses
All data are presented as the mean±standard error (SE). All statistical analyses were performed using GraphPad Prism (GraphPad Software Inc., La Jolla, CA, USA) and SigmaPlot software (ver. 12; Systat Software, San Jose, CA, USA). Statistical significance was determined using an unpaired 2-tailed Student's t-test and p<0.05 was considered significant.
RESULTS
Stimulation of splenocytes with silk peptide enhances NK cell activity
To preliminarily confirm the effects of silk peptide treatment on NK cell activation, mouse splenocytes were prepared and incubated with various concentrations of silk peptide. Silk peptide-stimulated splenocytes were then incubated with PKH-26-labeled YAC-1 target cells to measure NK cell-mediated cytotoxicity against target cells (Fig. 1). When splenocytes stimulated with silk peptide for 48 h (Fig. 1A) or 72 h (Fig. 1B) were incubated with YAC-1 target cells at an effector to target ratio of 10:1, we detected significantly enhanced target cell lysis at all of the silk peptide concentrations tested. Although we did not use purified NK cells in this ex vivo experiment, we can assume that treatment of splenocytes with silk peptide activates NK cells and wanted to confirm this effect in vivo.
 | Figure 1Effect of ex vivo silk peptide treatment of splenocytes on NK cell cytolytic activity against YAC-1 target cells. Splenocytes were incubated with the indicated concentrations of silk peptide for (A) 48 h or (B) 72 h and incubated with PKH-26-labeled YAC-1 target cells at a ratio of 10:1. The degree of target cell lysis was measured as described in the Materials and Methods. Data are presented as the mean±SE (n=3) of three independent experiments.E:T, effector to target ratio.
***p<0.001, ****p<0.0001 indicate a significant difference compared to the control.
|
Silk peptide enhances the proliferation of pre-stimulated splenocytes
To understand the effect of silk peptide treatment on the stimulation of immune cells, including NK cells, in vivo, mice were orally administered various concentrations of silk peptide-supplemented feed for 2 months. Initially, we determined whether oral silk peptide treatment influenced immune organs such as the spleen and thymus. There was no significant change in spleen or thymus weight, even with the highest amount of silk peptide (7,500 mg/kg body weight/day) administered (data not shown).
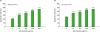
We next determined whether oral administration of silk peptide affects mitogen- or IL-2-stimulated proliferation of splenocytes (Fig. 2). The proliferation of mitogen- and IL-2-stimulated splenic lymphocytes was significantly increased in silk peptide-treated mice compared to control mice. For example, splenocytes prepared from high doses of silk peptide (3,000 and 7,500 mg/kg body weight/day)-treated mice prominently responded to Con A and LPS treatment by approximately 3- to 6-fold, respectively (Fig. 2A and B), relative to the control. Splenocytes simultaneously prepared from the intermediate dose group (1,500 mg/kg body weight/day) showed the highest proliferative response by IL-2 treatment (approximately 6-fold compared to the control) (Fig. 2C). Collectively, we conclude that oral silk peptide treatment in mice primes lymphocytes to stimulants, including mitogens and cytokine.
 | Figure 2Influence of oral administration of silk peptide on mitogen- or IL-2-mediated proliferation of splenocytes. Mice were orally administered various amounts of silk peptide-supplemented feed for 2 months and splenocytes were prepared. The level of (A) Con A-, (B) LPS-, or (C) IL-2-mediated proliferation of splenocytes was measured as described in the Materials and Methods. Data are presented as the mean±SE (n=3) of three independent experiments.
*p<0.05, **p<0.01, ***p<0.001, and ****p<0.0001 indicate a significant difference compared to the control (without silk peptide treatment).
|
Oral administration silk peptide in mice increases NK cell frequency and maturation
We next determined whether oral silk peptide treatment affects the subpopulation frequency in splenic lymphocytes. Initially, we analyzed changes in the CD4/CD8 cell ratio in mouse splenocytes and found that oral administration of silk peptide slightly reduced the CD4/CD8 cell ratio in splenocytes compared to the control (data not shown). We next examined the changes in NK cell frequency in mice orally fed silk peptide (Fig. 3A). We detected a significant increase of NK cell frequency in mice treated with the intermediate dose of silk peptide (p<0.05 and 0.01 in 750 and 1,500 mg/kg body weight/day-treated groups, respectively) compared to the control. We also analyzed the maturation status of NK cells following oral administration of various amounts of silk peptide (Fig. 3B). Mouse NK cells were subdivided into four subsets based on the expression levels of CD27 and CD11b on their surface as follows (naïve to mature phenotype): CD11blowCD27low, CD11blowCD27high, CD11bhighCD27high, CD11bhighCD27low (2930). The proportion of mature NK cells was increased by oral administration of silk peptide in a dose-dependent manner, and the enhancement was statistically significant (p<0.05 and 0.01) in mice fed high doses (3,000 and 7,500 mg/kg body weight/day, respectively) of silk peptide, where NK cell frequency was unchanged or slightly reduced compared with control mice (Fig. 3A). As a result, we suggest that oral administration of low to intermediate doses of silk peptide in mice induces the proliferation of NK cells and that oral administration of high doses of silk peptide may induce the maturation of NK cells.
 | Figure 3Influence of oral administration of silk peptide on NK cell frequency and the status of NK cell maturation in splenocytes. Changes in (A) NK cell frequency (CD3−NK1.1+) and (B) mature NK cells (CD11bhighCD27low) in splenocytes prepared from mice orally administered various amount of silk peptide were analyzed as described in the Materials and Methods. FACS plot shows a representative result and data shown in bar graph are presented as the mean±SE (n=3) of three independent experiments.FSC-A, forward-scatter area; SSC-A, side-scatter area.
*p<0.05 and **p<0.01 indicate a significant difference compared to the control (without silk peptide treatment).
|
Oral administration of silk peptide enhances NK cell-mediated target cell lysis
Given that oral administration of silk peptide in mice enhanced NK cell frequency and maturation, we were interested in the functional activation of NK cells following oral silk peptide administration. NK cells were purified from splenocytes prepared from mice following oral administration of various amounts of silk peptide for 2 months. Cells were then incubated with YAC-1 target cells at an effector to target ratio of 5:1. When the frequency of cells expressing CD107a, a functional marker of NK cells excreting granules, was examined among NK1.1+ cells, the CD107a+ cell frequency increased in a dose-dependent manner and the frequency was significantly higher than that of the negative control at all silk peptide doses tested (Fig. 4A). We also confirmed the functional activity of NK cells purified from silk peptide-treated mice by determining the frequency of IFN-γ-expressing cells, because IFN-γ is the most critical cytokine in NK cell activity (Fig. 4B). Similar to CD107a+ cell frequency, the number of IFN-γ-expressing cells also increased in a dose-dependent manner and was significantly enhanced in most doses of silk peptide-treated mice (Fig. 4B).
 | Figure 4Cytolytic activity of NK cells purified from splenocytes. NK cells were purified from splenocytes prepared from mice fed various amounts of silk peptide and stimulated with YAC-1 target cells at a ratio of 5:1 in the presence of brefeldin A and monensin. NK cell cytolytic activity was determined by measuring (A) cell surface CD107a expression and (B) intracellular IFN-γ expression as described in the Materials and Methods. FACS plot shows a representative result and data shown in bar graph are presented as the mean±SE (n=3) of three independent experiments. NC and PC show the minimum and maximum levels, respectively.FSC-A, forward-scatter area; SSC-A, side-scatter area; NC, negative control (unstimulated control containing NK cell only without target cells); PC, positive control (NK cells stimulated with PMA and ionomycin).
*p<0.05 and **p<0.01 indicate a significant difference compared to the control (without silk peptide treatment).
|
Silk peptide treatment stimulates the expression of cytokines that impact Th1-type immune responses
To analyze the pattern of immune modulation induced by oral administration of silk peptide in mice, cytokine expression characteristics of splenocytes prepared from silk peptide-treated mice were examined following in vitro stimulation with PMA together with ionomycin (Fig. 5). The expression of Th1-type cytokines, such as IFN-γ and IL-2, was significantly enhanced in splenocytes prepared from silk peptide-treated mice, although the enhancement varied depending on the dose of silk peptide (Fig. 5A and B). By contrast, the expression levels of Th2-type cytokines, such as IL-4 and IL-5 (not detected), were not significantly affected by oral administration of silk peptide (Fig. 5C). Consequently, we conclude that oral administration of silk peptide primes Th1-type immune responses, which include cell-mediated immune functions such as NK cell activity.
 | Figure 5Level of cytokine expression following stimulation of splenocytes prepared from mice fed various amount of silk peptide with PMA and ionomycin. Splenocytes prepared from silk peptide-treated mice were stimulated in vitro with PMA and ionomycin. The expression levels of the Th1-type cytokines (A) IFN-γ and (B) IL-2, and the Th2-type cytokine (C) IL-4 were determined as described in the Materials and Methods. Data are presented as the mean±SE (n=3) of three independent experiments.
*p<0.05 indicates a significant difference compared to the control (without silk peptide treatment).
|
DISCUSSION
Proper immune regulation is important for disease prevention and treatment in humans. Numerous research reports claim that decreased immune function weakens resistance to illness and healthy food consumption strengthens the immune system. In particular, it has been reported that certain protein food products, including lactoferrin, soybean protein fraction (a milk-derived protein casein), contain immune-modulating functions and thus increase NK cell activity in immunocompromised mice, stimulate the cellular immune response, and exert mitogenic effects (31323334). Silk protein has also been proposed as a bioactive substance and is effective at reducing blood pressure, exhibiting anti-tumor activity, treating atopic dermatitis, promoting insulin release, and enhancing innate immune responses (1418353637). In this regard, we explored the effects of silk peptide on immune capability in vivo, especially on NK cell activity, in mice.
The thymus is a central lymphoid organ of cell-mediated immunity in which T lymphocyte maturation occurs. In addition, the spleen is a peripheral lymphoid organ and plays an important role in triggering specific immune responses to antigens in the bloodstream. Therefore, an increase in the weights of the thymus and spleen is recognized as the induction of non-specific and specific immune response (38). Interestingly, the weights of the thymus and spleens from the silk peptide-treated group were not significantly different than the control (data not shown). We believe that silk peptide does not have direct mitogenic effects on lymphocytes per se, which was confirmed in vitro (manuscript in preparation). However, the proliferation of mitogen- and IL-2-stimulated lymphocytes from silk peptide-treated mice was significantly enhanced compared to control mice (Fig. 2). Consequently, we believe that the proliferation capability of immune cells associated with induction of the immune response following exposure to external antigens and stimuli is increased by oral administration of silk peptide, which should improve induction of the defense immune response.
As an immune cell that defends against viral infection in the early stages, NK cells play an important role in the selective elimination of irregular cells, such as cancer cells or infectious cells, by directly attacking and killing target cells. In healthy individuals, tumors and cancer cells are primarily removed by NK cells, whereas the frequency and function of NK cells are greatly reduced in cancer patients. Increasing the number and activity of NK cells would not only substantially improve the immune system of cancer patients, but also effectively help treat cancer patients. Therefore, a treatment method that increases immunity and the number of NK cells is urgently needed. Various methods for activating and promoting the proliferation of NK cells to increase anti-cancer immunity have been studied. NK cell-based cures have been promising and groundbreaking and have been clinically tested in customized chemotherapy. In cell-mediated immunotherapy, an important goal is obtaining a large number of NK cells by efficiently proliferating NK cells (3940). Although oral administration of high doses of silk peptide inhibited NK cell frequency, oral administration of intermediate doses (750 and 1,500 mg/kg body weight/day) significantly increased the frequency of NK cells (Fig. 3A). Interestingly, oral administration of silk peptide enhanced the frequency of mature NK cells in a dose-dependent manner, and even high doses of silk peptide, which inhibited NK cell frequency, significantly enhanced NK cell maturation (Fig. 3B). Exocytosis of the intracellular granule accompanies the cell surface expression of CD107a, and we confirmed that the ratio of CD107a-expressing cells increases in a dose-dependent manner to a statistically significant level following oral administration of the silk peptide in mice (Fig. 4A). We also confirmed that oral administration of silk peptide in mice increased the frequency of NK cells expressing intracellular IFN-γ, the most important cytokine for NK cell function (41) (Fig. 4B).
The present study showed that oral administration of silk peptide stimulates NK cell to proliferate and/or mature depending on the dose of silk peptide administered. The same treatment also activates NK cell cytotoxicity. As such, we believe that oral consumption of silk peptide is an effective way to induce the proliferation and activation of NK cells. We are currently testing the effect of silk peptide by using purified NK cells to confirm if silk peptide-mediated enhancement on NK cell activity is direct or indirect. Intake of silk peptide will also stimulate NK cells to influence systemic immune functions to improve natural immunity or may be used as a complementary therapy to support optimal NK cell activity in cancer patients.
 XML Download
XML Download