

 PDF
PDF Citation
Citation Print
Print


Abbreviations
AhR
aryl hydrocarbon receptor
APC
antigen presenting cell
BM
bone marrow
BSA
bovine serum albumin
DC
dendritic cell
DNFB
dinitrofluorobenzene
DTH
delayed type hypersensitivity
FICZ
6-formylindolo[3,2-b]carbazole
Flt3L
FMS-like tyrosine kinase 3 ligand
I3C
indole-3-carbinol
I3S
indoxyl 3-sulfate
IDO-1
indoleamine 2,3-dioxygenase-1
IFN
interferon
LBD
ligand binding domain
MEF
mouse embryonic fibroblast
NF-κB
nuclear factor B
pDC
plasmacytoid dendritic cell
PBS
phosphate buffered saline
STAT3
signal transducer and activator of transcription 3
TCDD
2,3,7,8-tetrachlorodibenzo-p-dioxin
TGF
transforming growth factor
Th
T helper
TLR
toll-like receptor
Treg
regulatory T cell
VSV
vesicular stomatitis virus
INTRODUCTION
Dendritic cells (DCs), of which morphology is characterized by dendrite-like extensions, play a central role in antigen recognition, activation of immediate and long-term immunity, and maintaining tolerance to self-antigens (12). DCs, which are heterogenous in terms of cell-surface markers, genetic program, anatomic location, and abilities to regulate T-cell fate, can be divided into 3 major populations in mice: 1) nonlymphoid tissue migratory DCs (also termed tissue DCs), 2) lymphoid tissue-resident DCs (also termed conventional DCs, and 3) plasmacytoid dendritic cells (pDCs) (3).
The pDCs was first identified as a cell type similar to plasma cells but lacking B cell and plasama cell markers (4). The pDCs secrete large amounts of type I interferons (IFNs) in response to viral antigens through Toll-like receptor (TLR) 7 and TLR9 (5), and pDCs can also secrete other pro-inflammatory cytokines and chemokines, including IL-12, CXC-chemokine ligand 8 and 10, promoting the polarization of CD4+ T cells into T helper (Th) 1 cells (6). In addition, pDCs secrete transforming growth factor (TGF)-β and express indoleamine 2,3-dioxygenase, promoting regulatory T cell (Treg) commitment. Unlike migratory DCs and tissue-resident DCs which are inherently efficient antigen-presenting cells (APCs) capable of eliciting lymphocyte responses, pDCs are considered to be poor APCs based on their low surface expression of major histocompatibility complex and costimulatory molecules (7). However, pDCs can become potent APCs capable of activating T cells upon stimulation with CD154 and TLRs (8). Thus, in addition to contributing innate immunity, pDCs can be immunogenic and tolerogenic.
The aryl hydrocarbon receptor (AhR) is a ligand-activated transcription factor that belongs to the basic region-helix-loop-helix sub-family of DNA binding proteins with Per-Arnt-Sim domains (9). AhR is a cytosolic sensor of small synthetic and natural chemicals, which act as its ligands. Upon binding of a ligand to AhR, it undergoes a conformational change for translocation to the nucleus where it forms a heterodimeric complex with the Ah receptor nuclear translocator (10). This complex activates transcription of target genes such as Cyp1A1 and glutathione S-transferase by binding to the aromatic hydrocarbon-responsive elements in the 5′ flanking region of these genes (11). Environmental AhR ligands are exposed to the barrier organs such as the skin, lung and gut, with the gut accounting for most of human dioxin exposure and for other AhR ligands, including many dietary components such as quercetin in apples, resveratrol in red wine and indole-3-carbinol (I3C) in many cruciferous plants (1213).
AhR, which is constitutively expressed in DCs (14), regulates the activation, maturation and differentiation of DCs. AhR activated bone marrow (BM)-derived DCs and was involved in Langerhans cell maturation (1516). Effects of AhR on the differentiation of DCs are different depending on cell types used. In 1 study, AhR activation promoted human monocyte-derived DC differentiation (17), whereas in other studies, AhR inhibited in vitro differentiation of Langerhans DCs and myeloid DCs from CD34+ hematopoietic progenitor cells (1819). Of interest, accumulating evidence indicates that AhR activation induces immune tolerance via a DC-mediated mechanism (202122). AhR agonist VAF347 promoted in vivo allograft tolerance via DC-mediated effects on Tregs (20). The same compound exerted anti-inflammatory effects by inhibiting the production of inflammatory cytokines and the upregulation of costimulatory molecules on human monocyte-derived DCs (21). Activation of AhR by 2-(1′H-indole-3′-carbonyl)-thiazole-4-carboxylic acid methyl ester, an endogenous ligand, induced not only FoxP3+ Treg that suppressed experimental autoimmune encephalomyelitis, but also tolerogenic DCs (22).
Although it is still not clear whether the regulatory functions of DCs is determined by their activation status or inherent of different lineages, in vitro and in vivo experiments have identified several subsets of DCs of tolerogenic characteristics (23): 1) CD11c+ DCs expressing perforin that enforces peripheral tolerance by deleting T cells, 2) CD103+ DCs present in the intestinal mucosa, where they play a key role in oral tolerance, and 3) pDCs.
We previously reported that AhR ligand 3,3′-diindolylmethane, an acid-stimulated conversion product of I3C, inhibited FMS-like tyrosine kinase 3 ligand (Flt3L)/granulocyte-macrophage colony-stimulating factor-induced BM-derived CD103+ DC differentiation (24). AhR antagonist StemRegenin 1 promoted human pDC development from CD34+ hematopoietic progenitor cells (19). In mice, lack of AhR promoted pDC development in vivo and AhR activation by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) inhibited pDC differentiation from BM in vitro (25). However, the underlying inhibitory mechanisms of AhR in pDC development and the physiological significance of AhR activation have not been addressed.
Since the first report that AhR is the cytosolic receptor for TCDD (26), a number of low-molecular weight, structurally diverse chemicals, including metabolites of tryptophan and arachidonic acid, indoles, and tetrapyroles, have been identified as naturally occurring exogenous and endogenous AhR ligands (27). Interestingly, although all AhR ligands bind AhR activating AhR, AhR functionality often depends on ligands used, showing ligand-selective effects on cell proliferation, death, and differentiation, due to differential affinity of AhR for structurally diverse ligands, stability of ligands, and differential binding of AhR ligands within the AhR ligand binding domain (28). TCDD, which is metabolically stable, induced Treg which suppressed experimental autoimmune encephalomyelitis, whereas 6-formylindolo[3,2-b]carbazole (FICZ), a photoproduct generated from tryptophan by ultraviolet B irradiation and an endogenous AhR ligand which is metabolically unstable (29), interfered with Treg development, increasing the severity of experimental autoimmune encephalomyelitis (30). In a mouse model of influenza virus infection, TCDD modulated inflammatory responses characterized by neutrophilia and virus-specific CD8 T cell response, whereas FICZ had no effect to infection (31). In various species, TCDD shows a spectrum of AhR-dependent toxic effects (i.e., wasting, dermal toxicity, thymic involution, and teratogenicity), which are not observed with nonhalogenated polycyclic aromatic hydrocarbons (28). Thus, interpretation of results studied with TCDD needs caution.
In the present study, using in vitro pDC differentiation model in which BM cells are cultured with FLt3L for 9 days, we examined effects of 2 AhR ligands, indoxyl 3-sulfate (I3S), a uremic toxin that originates from the metabolism of tryptophan (32), and I3C, a phytochemical that is abundant in cruciferous plants, on pDC differentiation and investigated underlying molecular mechanisms. Finally, using a mouse model of dinitrofluorobenzene (DNFB)-specific skin delayed type hypersensitivity (DTH), whether oral tolerance to DNFB is modulated by AhR activation was addressed.
MATERIALS AND METHODS
Mice
C57BL/6 mice, which were purchased from the Central Lab. Animal Inc. (Seoul, Korea), were used at the age of 6–12 weeks. The animals were housed 5 mice per cage in a laminar air flow room maintained at 22°C±2°C with relative humidity of 55%±5%. Mice were cared and treated in accordance with the guidelines established by the Changwon National University public health service policy on the use of laboratory animals.
Chemicals and reagents
I3S, I3C, and DNFB were purchased from Sigma-Aldrich (St. Louis, MO, USA); murine Flt3L from eBioscience (San Diego, CA, USA). Antibodies used in the present study are: anti-signal transducer and activator of transcription 3 (STAT3) from Santa Cruz Biotechnology (Santa Cruz, CA, USA); anti-phospho-c-Src (Tyr416) from Cell Signaling Technology (Danvers, MA, USA); anti-phospho-STAT3 (Tyr705) from BD Biosciences (Franklin Lakes, NJ, USA); FITC anti-mouse CD11c, Alexa Fluor 647 anti-mouse B220, anti-ERK, and anti-phospho-ERK (Thr202/Tyr204) from Biolegend (San Diego, CA, USA). CpG oligodeoxyribonucleotides (CpG 2216: sequences are shown 5′-3′: small letters, phosphoorothioate linkage; capital letters, phosphodiester linkage 3′ of the base; bold, CpG-dinucleotides; ggGGGACGATCGTCgggggG) were manufactured by and purchased from Bioneer (Daejeon, Korea).
Development of CD11c+B220+ DC from BM cells
Development of CD11c+B220+ DC from BM cells was previously described (33). Briefly, BM cells were isolated by flushing femurs with RPMI 1640 (Roswell Park Memorial Institute, Buffalo, NY, USA) supplemented with 2.5 mM/L HEPES buffer (pH 7.4) and depleted of red blood cell by being treated in Tris-ammonium chloride at room temperature for 2 min. BM cells were cultured in a 24-well plate at a density of 1×106 cells/ml in RPMI 1640 with 10 mM HEPES, 2 mM L-glutamine, 10% fetal bovine serum, 50 μM β-mercaptoethanol, and Flt3L (200 ng/ml) for 9 days.
AhR ligand treatment
For in vitro experiments, AhR ligands dissolved in DMSO were added to the culture at the final concentration of 1 μM 1h before the addition of Flt3L. DMSO (0.1% v/v) was used as vector control. For gastric administration, 52 μg of I3S, 2 mg of I3C, or 10 μl of DMSO was mixed with 140 μl of corn oil and was given to mice at days 1, 3, 5, 7, and 9. At day 10, oral tolerance to DNFB was induced.
Flow cytometry and cell sorting
Cells were kept on ice or at 4°C at all time. Immunofluorescence staining was carried out in 96-well, round-bottomed cell culture plates. The cells were centrifuged and resuspended in 10 μl of primary antibody diluted with PBS at a previously determined optimal concentration. After 15 min of incubation, the cells were washed with washing buffer (PBS/0.5% bovine serum albumin (BSA)/0.09% sodium azide) 3 times and fixed with 0.9% buffered formalin. Cells were analyzed within 1 week on a FACSCalibur with CellQuest software (BD Biosciences). For cell sorting, BM cells cultured for 9 days in the presence of Flt3L were stained with FITC anti-mouse CD11c and Alexa Fluor 647 anti-mouse B220, and sorted with FACS Aria III. The cut-off point was the co-expression of CD11c and B220 at high levels.
IFN-α production assay
BM cells were incubated with FLt3L for 8 days and then treated with CpG oligodeoxyribonucleotides (CpG 2216) (3 μg/ml) for 24 h. Supernants were then collected and assayed by ELISA for murine IFN-α (MyBiosource, San Diego, CA, USA), according to the manufacturer's instructions. The limit of detection for IFN-α was 9 pg/ml.
RNA preparation and RT-PCR
Total cellular RNA was extracted from cells using the RNAzol method (Tel-Test Inc., Friendswood, TX, USA). For PCR analysis, RNA was used after contaminating DNA was completely removed by DNase I treatment. RT-PCR analysis was performed using pairs of oligonucleotide primers. The PCR products were confirmed to correspond to their original sequence by DNA sequencing. Gene specific primers, number of cycles of amplification, annealing temperature, and expected size of PCR product are listed in Table 1.
Table 1
Primers used in RT-PCR

![]()
Western blotting
Cells or tissues were homogenized in lysis buffer containing 20 mM Tris-HCl (pH 7.4), 1 mM EDTA (pH 8.0), 50 μM sodium vanadate, 20 mM p-nitrophenylphosphate, 50 mM sodium fluoride, leupeptin (0.5 μg/ml), aprotinin (10 μg/ml), and soybean trypsin inhibitor (10 μg/ml). Proteins size-fractionated on SDS/polyacrylamide gel electrophoresis were transferred to polyvinylidene difluoride membranes, and the blots were blocked with 3% BSA in tris-buffered saline (TBS) buffer (20 mM Tris-HCl pH 7.5/137 mM NaCl). The blots were sequentially treated with primary and secondary antibodies in TBS with Tween 20 (20 mM Tris-HCl pH 7.5/137 mM NaCl/0.1% Tween 20) with intermittent washing with TBS with Tween 20. Immunodetection was performed with the ECL-plus kit (Amersham Biosciences, Piscataway, NJ, USA).
Oral tolerance induction to DNFB
Oral tolerance to DNFB was induced by a single intragastric administration of 0.3 ml of 0.1% DNFB in acetone/olive oil (1/10, v/v) 7 days before skin sensitization with DNFB, as described (34).
Contact hypersensitivity to DNFB
Contact hypersensitivity to DNFB was determined by the mouse ear swelling test as described (34). Briefly, mice were sensitized epicutaneously onto 2 cm2 of shaved abdominal skin with 25 μl of 0.5% DNFB in acetone-olive oil (4:1, v/v) and challenged on day 5 with 4 μl of 0.2% DNFB, applied onto each side of the right ear. The left ear was painted with the vehicle alone. Ear thickness was measured using a digital caliper (Mitutoyo 500-197-30; Mitutoyo America Corporation, Aurora, IL, USA) before and after challenge. The ear swelling was calculated as (T-To of the right ear) – (T-To of the left ear), in which T and To represent the values of ear thickness after and before challenge, respectively.
RESULTS
Inhibition of pDC differentiation from BM cells by I3S and I3C
A large number of murine pDCs can be generated from BM precursors with Flt3L (35). Thus, BM cells were cultured for 9 days in the presence of Flt3L and then assayed for the expression of CD11c/B220, surface markers for murine pDCs (Fig. 1A). We performed this experiment 3 times. On average, about 6% of the harvested cells were CD11c+B220+. It is well known that hematopoietic stem cells respond to xenobiotics such as TCDD (36). To examine whether BM cells and CD11c+B220+ pDCs functionally express AhR, CD11c+B220+ cells were sorted using FACS Aria III, treated with I3S or I3C, and assayed for the expression of AhR and Cyp1A1, a target gene of AhR. AhR, which was expressed in BM was increased in Flt3L-treated BM cells and CD11c+B220+ cells (Fig. 1B). For Cyp1A1 expression, 3 days after BM cells were cultured in the presence of Flt3L with or without I3S and I3C, cells were harvested and analyzed for Cyp1A1 expression. Cyp1A1 was induced in Flt3L-treated BM cells by I3S or I3C (Fig. 1C), suggesting that AhR is functional in BM-derived DCs. To investigate the role of AhR activation in the differentiation of pDCs from BM in vitro, BM cells were cultured for 9 days in the presence of AhR ligands I3S or I3C. AhR activation by I3S and I3C showed almost no effect on the total number of cells generated from BM precursors (Fig. 1D). However, I3S and I3C significantly reduced the proportion of CD11c+B220+cells (5.2%, p=0.004 and 4.1%, p=0.001, respectively), compared with PBS and DMSO controls (6.6% and 7.1%, respectively) (Fig. 1E and F).
 | Figure 1I3S and I3C inhibit the development of CD11c+B220+ pDCs. BM cells were induced to differentiate into CD11c+B220+ DCs with Flt3L for 9 days. Then, cells were harvested and analyzed for cell surface expression of CD11c and B220 (A) mRNA expression of AhR and Cyp1A1 (B, C) and the effects of AhR on cell growth (D), and the proportion of CD11c+B220+ cells (E). For AhR expression in CD11c+B220+ pDCs, CD11c+B220+ cells, which were sorted from the 9-day old BM cultures, were subjected to RT-PCR (B). The results for the effects of AhR on the development of CD11c+B220+ cells were analyzed by the paired Student's t-test and presented as mean±standard deviation (n=3 experiments) (F).
*p<0.05 compared with DMSO.
|
Reduction of IFN-α expression in Flt3L-cultured BM cells treated with I3S and I3C
The pDCs secrete large amounts of type I IFNs in response to viral antigens through TLR9. Thus, we examined if AhR activation alters IFN-α/β producing ability of pDCs by treating BM cultures with CpG oligodeoxyribonucleotides (CpG 2216), a synthetic ligand for TLR9 (37). Twenty-four h after CpG administration, cells were harvested and assayed for IFN-α by RT-PCR and ELISA. IFN-α mRNA expression was reduced by I3S and I3C (Fig. 2A). Accordingly, the amount of IFN-α in the culture supernatant was also deceased in BM cultures treated with I3S and I3C, compared with PBS and DMSO controls (Fig. 2B).
 | Figure 2I3S and I3C downregulate IFN-α production from Flt3L-treated BM cultures. BM cells were induced to differentiate into CD11c+B220+ DCs with Flt3L for 8 days and activated with CpG oligodeoxyribonucleotides (CpG 2216) for 24 h. Then, cells were harvested and analyzed for mRNA expression of IFN-α by RT-PCR and supernatants were assayed for IFN-α by ELISA.
|
Downregulation of the expression of STAT3 and E2-2 by AhR activated with I3S and I3C
Flt3L play a key role in pDC development (38). Flt3L promotes pDC development by activating STAT3, which upregulates the expression of E2-2. E2-2 (Tcf4), which is the master transcription factor for pDC development, in turn regulates genes associated with function and development of pDC (6). Thus, we first investigated effects of AhR on the phosphorylation of STAT3 in BM cells treated with Flt3L for 1 h (Fig. 3A). Phosphorylation of STAT3, which was increased by Flt3L, was reduced by I3S or I3C. Whether the downregulation of STAT3 phosphorylation by I3S and I3C is due to transcriptional or post-transcriptional regulation, STAT3 was quantified. Interestingly, the level of STAT3 protein, which was unchanged by Flt3L, was decreased by I3S and I3C, suggesting that AhR activation downregulates the expression of STAT3. AhR also functions by interacting with other signals, altering activation of those signals such as receptor tyrosine kinases and mitogen-activated protein kinase (28). Thus, we examined if AhR activation by I3S and I3C regulates activation of ERK and c-Src. AhR showed little effect both on the activation of ERK and c-Src and the expression of ERK, suggesting that AhR works specifically on STAT3. Next, we investigated effects of AhR on the expression of transcription factors required for pDC development in BM cells treated with Flt3L for 9 days (Fig. 3B). Expression of PU1, IRF8, and E2-2 was increased in Flt3L-treated BM cells compared with BM cells. Of the 3 genes, only E2-2 expression was downregulated by I3S and I3C. pDC promotes Treg differentiation by expressing indoleamine 2,3-dioxygenase-1 (IDO-1) and TGF-β (6). Thus, effects of AhR activation on immunosuppressive functions of pDC were examined in BM cells treated with Flt3L for 9 days. Expression of IDO-1 and TGF-β, which was increased by Flt3L treatment, was reduced by I3S and I3C (Fig. 3C), suggesting that AhR activation by I3S and I3C inhibits immunosuppressive function of pDC.
 | Figure 3I3S and I3C regulates expression of genes associated with development and functions of pDCs. BM cells, which were cultured in the presence of Flt3L for 1 h, were harvested and analyzed by western blotting (A) or RT-PCR (B). For analysis of genes associated with pDC functions, BM cells cultured supplemented with Flt3L for 9 days were analyzed by RT-PCR.
|
Impairment of oral tolerance to DNFB in mice administrated with I3S or I3C accompanied by a decrease in the proportion of CD11c+B220+ cells in mesenteric lymph nodes
Oral tolerance, which is the immunological mechanism by which the mucosal immune system protects the host from deleterious T-cell mediated reactions to self and exogenous dietary and environmental antigens, is mainly mediated by IL-10 and/or TGF-β-producing Tregs (39). Treg development in the mucosa is promoted by tolerogenic DCs, including CD103+ DCs and pDCs (4041). Effects of AhR on oral tolerance seem to be bipolar, depending on ligands used. In studies of the induction of oral tolerance against ovalbumin, TCDD impaired oral tolerance whereas I3C supported oral tolerance (4243). Thus, we examined if oral tolerance is affected by AhR activated by I3S and I3C. Mice, which were fed I3S and I3C and then DNFB 7 days before skin sensitization with DNFB, were ear-challenged with DNFB 5 days later. Contact hypersensitivity was determined by the mouse ear swelling test (Fig. 4A). Control mice orally injected with PBS developed ear swelling response which lasted several days. Oral injection of DMSO (vehicle) inhibited contact hypersensitivity to DNFB. When DNFB was co-injected with I3S or I3C, contact hypersensitivity to DNFB was not suppressed, suggesting that induction of oral tolerance to DNFB is suppressed by AhR activated by I3S and I3C. One type of tolerogenic DCs in the mucosal immune system is pDC. Thus, we sacrificed mice on the fourth day after the ear-challenging of DNFB, collected mesenteric lymph nodes, and analyzed total cells for the expression of CD11c/B220. In mice treated with I3S and I3C, the proportion of CD11c+B220+ cells was greatly reduced (0.62%, p=0.00009 and 0.69%, p=0.00009, respectively), compared with PBS oral or DMSO control (1.32% and 1.15%, respectively) (Fig. 4B and C).
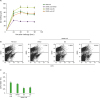 | Figure 4I3S and I3C impair oral tolerance to DNFB and reduce the proportion of CD11c+B220+ cells in the mesenteric lymph nodes. Mice, which were fed I3S or I3C, were given a single intragastric administration of 0.3 ml of 0.1% DNFB in acetone/olive oil (1/10, v/v). Seven days later, the mice were sensitized epicutaneously onto 2 cm2 of shaved abdominal skin with 25 μl of 0.5% DNFB in acetone-olive oil (4:1, v/v) and challenged on day 5 with 4 μl of 0.2% DNFB, applied onto each side of the right ear. Ear thickness was measured before and after challenge (A). Results of (A) are presented as mean±standard deviation (n=5 mice per group). At 96 h after the challenge of DNFB, mesenteric lymph nodes were collected, stained with flurochrome-conjugated ant-CD11 Ab and anti-B220 Ab, and analyzed for expression of CD11c and B220 (B). Results of (B) are statistically analyzed and presented as mean±standard deviation (n=5 mice per group) in (C).
*p<0.05 compared with DMSO.
|
DISCUSSION
This study demonstrates that AhR ligands I3S and I3C inhibit the development of pDCs from BM cells stimulated with Flt3L in vitro, downregulating the expression of STAT3 and E2-2. Intragastric administration of I3S and I3C abrogated oral tolerance to DNFB characterized by suppression of DTH responses to DNFB and reduced the proportion of CD11c+B220+ cells in the mesenteric lymph nodes.
In the present study, we observed that phosphorylation of STAT3, which was enhanced by the addition of Fl3tL, was inhibited by I3S and I3C. However, expression and activation of ERK and activation of c-Src were found not to be modulated by I3S and I3C, suggesting that effects of I3S and I3C on signaling are specific on STAT3. Interestingly, the level of STAT3 protein itself was reduced by AhR activation, suggesting that AhR could regulate the expression of STAT3, resulting in a proportional decrease of the phosphorylation of STAT3. There are several potential underlyng mechanisms for the downregulated expression of STAT3 protein. One possibility is the ubiquitin-proteasome system-mediated protein degradation. AhR is a ligand-dependent E3 ubiquitin ligase, degrading target proteins, including estrogen receptor-α and STAT1 and STAT4, as early as 1 h after AhR activation (4445). In the present study, BM cells were treated with AhR ligands 1 h before the addition of Flt3L, supporting the role of the ubiquitin-proteasome system in STAT3 degradation. Alternatively, AhR, which usually functions as a transcriptional activator, also suppresses gene expression (464748). Although AhR repressor, which is upregulated by AhR signaling, functions as a negative feedback regulator of AhR signaling (49), it could regulate gene expression by working as a transcriptional repressor of genes including peroxisome proliferator-activated receptor γ (48) or by interacting with other signaling pathways such as nuclear factor κB (NF-κB) (47). AhR also regulates gene expression by interacting with other signals including NF-κB and c-Src (2850). How STAT3 expression is regulated by AhR activation remains to be investigated.
In the present study, we demonstrated that oral tolerance induced by gastric administration of DNFB was impaired by I3S and I3C. In other studies, effects of AhR on oral tolerance were variable, depending on AhR ligands used; TCDD impaired oral tolerance whereas I3C supported it (4243). Furthermore, I3C showed different effects on oral tolerance induced by different antigens. The model of contact hypersensitivity to DNFB used to study effects of AhR on oral tolerance in the present study is mediated by effector CD8+ T cells (51) and CD4+CD25+ Tregs play a major role in the induction of oral tolerance to DNFB (39). In an oral tolerance model to ovalbumin, oral tolerance was determined by the level of serum antibodies to ovalbumin (43). In studies related to effects of AhR on CD4 T cell differentiation, activation of AhR by I3S promotes Th17 differentiation, while inhibiting Th2 differentiation and having little effect on Th1 differentiation, suggesting that AhR functions in a lineage-specific manner (5253). AhR can bind and be activated or inhibited by a wide variety of structurally dissimilar compounds via its ligand binding domain (LBD) (28). Differential binding of structurally diverse ligands within the AhR LBD could lead to ligand-dependent differences in the overall structure of the activated AhR, which may contribute to its ligand-dependent functional specificity by differentially recruiting coactivator or repressor and thus modulating transcriptional activity. AhR signaling could also be regulated by proteosomal degradation and AhR repressor. The AhR protein is rapidly downregulated following ligand binding, which occurs via ubiquitin-mediated 26S proteasome pathway (54). Interestingly, the kinetics of AhR degradation are different among ligands used (5556). Depletion of AhR persisted for at least 72 h after TCDD exposure, whereas treatment with 3-methylcholanthrane or β-napthoflavone caused a transient drop of AhR protein followed by a recovery of AhR to near pretreatment levels within 72 h, suggesting that the level of AhR protein remains downregulated when cells are treated with a metabolically stable ligand such as TCDD. AhR repressor, which regulates AhR signaling by acting as a transcriptional repressor (49), is transcriptionally induced by AhR activated by various ligands, including TCDD, 3-methylcholeanthrene, and benzo[a]pyrene. However, the degree of induction is variable depending on cell types and ligands used (5758). How AhR functions in a ligand-specific way remains to be investigated.
In the present study, we demonstrated that the population of CD11c+ B220+ pDCs in mesenteric lymph nodes of oral tolerance-induced mice was reduced by II3S or I3C. The induction and maintenance of oral tolerance is largely mediated by the combination of commensals, Treg, and tolerogenic DCs (59). In addition, CX3CR1+ macrophages, IL-22 producing innate lymphoid cells, and intestinal epithelial cells are also associated with oral tolerance induction (606162). Interestingly, these cells and CD103+ DCs also express the AhR protein (13246364). Thus, it remains to be tested whether AhR impairs oral tolerance by regulating the differentiation and function of those cells.
AhR activated by TCDD reduced survival rate of mice infected with influenza A virus (65) and viral titers in AhR-deficient mouse embryonic fibroblasts (MEFs) after infection with vesicular stomatitis virus (VSV) and influenza A virus increased (66). In mice exposed to TCDD and infected with influenza A virus, the proliferation and differentiation of CD8+ cytotoxic T cells were suppressed, which could be mediated by DCs regulated by AhR (6567). IFN-β production in AhR-deficient MEF after infection with VSV virus and influenza A virus increased compared with wild-type MEF and intraperitoneal pretreatment with FICZ resulted in a marked reduction in serum IFN-β protein at 12 h after VSV infection (66). How AhR regulates IFN-β production was elucidated. AhR upregulates expression of the ADP-ribosylase TCDD-inducible poly(ADP-ribose)polymerase, which in turn caused the downregulation of type 1 IFN response. In the present study, we demonstrated that AhR activated by I3S and I3C inhibited the differentiation of type 1 IFN-producing pDCs, suggesting that AhR could suppress the innate immune response against viral infection by regulating the development of DCs.
 XML Download
XML Download