

 PDF
PDF Citation
Citation Print
Print


Abbreviations
Arg-1
arginase-1
BMDM
bone marrow-derived macrophages
FBS
fetal bovine serum
GM-CSF
granulocyte macrophage colony-stimulating factor
IFN
interferon
iNOS
inducible nitric oxide synthase
IRF
interferon regulatory factor
KO
knockout
LPS
lipopolysaccharides
M-CSF
macrophage colony-stimulating factor
MHC
major histocompatibility complex
SEM
standard error of the mean
TGF
transforming growth factor
TNF
tumor necrosis factor
WT
wild type
INTRODUCTION
Macrophages play a critical role in antigen presentation, phagocytosis, and immunomodulation (1). They can be polarized into classically activated macrophages (M1) or alternatively activated macrophages (M2) in response to different stimulation (2). M1 and M2 macrophages have different roles in inflammation. M1 macrophages are associated with production of pro-inflammatory cytokines such as tumor necrosis factor (TNF)-α, IL-6, and IL-12 and provide a host defense against infections, while M2 macrophages secrete anti-inflammatory cytokines such as transforming growth factor (TGF)-β and IL-10 and have reparative function (34). M1 macrophages stimulated by interferon (IFN)-γ and/or lipopolysaccharides (LPS) or microbial product express high levels of CD68 marker, CD80 and CD86 costimulatory molecules, major histocompatibility complex (MHC) class II, and inducible nitric oxide synthase (iNOS) (34). M2 macrophages induced by IL-4 or IL-13 express increased levels of arginase-1 (Arg-1), CD206 mannose receptor, and CD163 marker (34). Moreover, recent studies showed that M1 and M2 macrophages can be converted into each other in their specific microenvironment (567).
Viperin (virus inhibitory protein, endoplasmic reticulum-associated, interferon-inducible) was first identified as a gene induced by IFN-γ in human primary macrophages and by human cytomegalovirus infection in human fibroblasts (89). It is also induced in a variety of cells by type I, II, and III IFNs, double-stranded (ds) B-form DNA, the dsRNA analog polyinosinic-polycytidylic acid, LPS, and many other viral infections (10). Viperin is a multifunctional protein. It functions as an anti-viral protein against many viruses in the majority of cell types including fibroblasts, hepatocytes, and astrocytes (811121314). This protein also plays a role as a regulator of cell signaling pathways in plasmacytoid dendritic cells and T helper 2 cells (1516). Moreover, it functions as a pro-viral protein by modulating cellular energy and lipid metabolism in fibroblasts infected with human cytomegalovirus (1718). Although viperin's functions in a variety of cell types were well known, the role of viperin in macrophages in which it was identified remains to be elucidated.
In this study, we showed that viperin is expressed in murine bone marrow cells including monocytes, and its expression is kept in bone marrow-derived macrophages (BMDMs) differentiated with macrophage colony-stimulating factor (M-CSF). Using BMDMs prepared from viperin knockout (KO) mice, we found that viperin deficiency facilitates polarization of BMDMs into M1 and M2 macrophages and secretion of cytokines, suggesting that viperin has a pivotal role as a regulator in macrophage polarization.
MATERIALS AND METHODS
Cells, antibodies, and reagents
Murine bone marrow cells used in the study were isolated from wild type (WT) or viperin (also known as Rsad2) KO C57BL/6 mice (18).
Viperin was detected with the monoclonal mouse antibody to viperin (MaP.VIP) described previously (813). The monoclonal mouse anti-β-actin antibody (Sigma-Aldrich, St. Louis, MO, USA), horseradish peroxidase-conjugated goat anti-rabbit or anti-mouse IgG antibody (Jackson ImmunoResearch Laboratories, West Grove, PA, USA), PE-conjugated anti-Ly-6C or anti-F4/80 antibody (BioLegend, San Diego, CA, USA), and PerCP-Cy5.5-conjugated anti-CD11b antibody (BD Biosciences, Franklin Lakes, NJ, USA) were used.
Mouse M-CSF or granulocyte macrophage colony-stimulating factor (GM-CSF) is stably expressed in CHO cells as described previously (19). The culture supernatants containing the secreted M-CSF or GM-CSF were used for generation of BMDMs.
Polarization of M1 and M2 macrophages
To generate BMDMs, the bone marrow cells from femurs and tibias of mice were harvested and cultured in RPMI (Hyclone™; GE Healthcare, Chicago, IL, USA) supplemented with 10% fetal bovine serum (FBS) (Hyclone™; GE Healthcare), 1% penicillin/streptomycin (Sigma-Aldrich), and 10% M-CSF or GM-CSF supernatants for 7 days. BMDMs cultured in M-CSF conditioned media were washed and plated at a density of 5–6 × 106 cells/well in 6-well plate (SPL Life Sciences, Pocheon, Korea) containing RPMI media without M-CSF. For M1 polarization, cells were classically activated with 100 ng/ml LPS (Sigma-Aldrich) + 20 ng/ml IFN-γ (R&D Systems, Minneapolis, MN, USA) for 24 h. For M2 polarization, cells were alternatively activated with 20 ng/ml IL-4 (PrimeGene, Shanghai, China) for 48 h.
Flow cytometry
BMDMs, M1 or M2 macrophages were blocked with 2.4G2 blocking buffer for 15 min at 4°C. Then, cells were surface stained with fluorescently conjugated FACS antibodies including V450-conjugated anti-CD86, FITC-conjugated anti-MHC II, and FITC-conjugated anti-CD206 antibodies and isotype control antibodies for 30 min at 4°C in dark condition. Cells were washed with 150 ul FACS buffer (1% FBS in PBS) and centrifuged at 2,000 rpm for 2 min. After washing with FACS buffer, single cell suspensions were obtained with cell strainers and resuspended in FACS buffer. Then cells were run on a LSR-II Fortessa Flow Cytometer (BD Biosciences). Data were analyzed with FlowJo software.
Cytometric bead array
To measure the levels of M1 and M2 secrete cytokines, the mouse TNF-α, IL-6, and IL-10 flex kits (BD Biosciences) were used. The assays were performed according to the manufacturer's instructions. Samples and standards were processed on the BD FACSVerse Flow Cytometer (BD Biosciences). Data were analyzed with FlowJo software (FlowJo, LLC, Ashland, OR, USA).
Immunoblot analysis
Cells were harvested and lysed in lysis buffer (1% Triton X-100 in TBS). The concentrations of protein were measured by bicinchoninic acid assay (Thermo Fisher Scientific, Waltham, MA, USA). The proteins were separated by 10% SDS-polyacrylamide gel electrophoresis gels and transferred to polyvinylidene difluoride membranes. The blots were blocked in 5% skim milk and 0.05% Tween in PBS for 1 h, incubated with primary antibodies, and probed with horseradish peroxidase-conjugated secondary antibodies, followed by incubation with enhanced chemiluminescence reagents (Thermo Fisher Scientific).
RNA extraction, cDNA preparation, and quantitative real-time PCR
Cells were harvested and total RNA were isolated using RNeasy Mini kit (Qiagen, Hilden, Germany). The cDNA synthesis was performed with 1 ug RNA using Prime script 1st strand cDNA synthesis kit according to the manufacturer's instructions (Takara Bio Inc., Kusatsu, Japan). The cDNA obtained from cells was quantified by quantitative real-time PCR using SYBR Green PCR kit (Applied Biosystems, Foster City, CA, USA). The reaction included 95°C for 10 min, which was followed by a 3-step PCR program of 95°C for 30 s, 55 °C for 1 min, and 72°C for 30 s repeated for 50 cycles. The PCR was performed in triplicate for each sample. Quantitation was performed by the comparative Ct (2−ΔΔCt) method. The Ct value for each sample was normalized by the value for β actin gene. Two independent experiments were analyzed statistically for differences in the mean values, and p values are indicated in the figures.
RESULTS
Viperin is basally expressed in murine bone marrow cells and BMDMs.
Viperin was identified in human primary macrophages treated with IFN-γ (8). To investigate the role of viperin in the macrophages, we isolated murine bone marrow cells from mice and assessed viperin expression in each cell population. Viperin expression was detected in most bone marrow cells including granulocytes (CD11bhighLy6Cmiddle, 19.1%), monocytes (CD11bmiddleLy6Chigh, 4.51%), and other cell populations (CD11blowLy6Clow; CD11bhighLy6Clow; CD11blowLy6Chigh) in the absence of IFN treatment (Fig. 1A). It has been well established that bone marrow cells differentiate into macrophages in M-CSF or GM-CSF conditioned media (202122). Viperin expression was maintained in the BMDMs (F4/80+CD11b+) differentiated with M-CSF but not with GM-CSF (Fig. 1B and C). Consistent with previous reports (813), viperin expression was significantly increased in BMDMs differentiated with M-CSF or GM-CSF in the presence of IFN and/or LPS treatment (Fig. 1C). The results indicated that viperin is basally expressed in murine bone marrow cells and its expression is maintained in BMDMs differentiated with M-CSF.
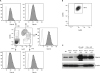 | Figure 1Viperin expression in murine bone marrow cells and BMDMs. (A) The expression of viperin in bone marrow cells. The murine bone marrow cells were isolated from WT C57BL/6 mice. The cells were stained with fluorescently-conjugated antibodies specific to CD11b, Ly6C, and viperin and analyzed by flow cytometry. Cell fractions were gated in a flow cytogram. Specific cell types were confirmed by expression levels of CD11b and Ly6C (Granulocytes, CD11bhighLy6Cmiddle; Monocytes, CD11bmiddleLy6Chigh). Viperin expression of each cell population was shown in flow histograms. Shaded histograms represent staining with anti-viperin antibody; open histograms represent staining with an isotype-matched control antibody of irrelevant specificity. Data are representative of two independent experiments. (B, C) The expression of viperin in BMDMs. The bone marrow cells were differentiated into BMDMs by incubation in M-CSF or GM-CSF conditioned media for 7 days. The cells were stained with fluorescently-conjugated antibodies specific to CD11b and F4/80. Cell fractions were analyzed by flow cytometry. BMDMs specificity was confirmed by expression of F4/80 and CD11b (F4/80+CD11b+). BMDMs were washed and plated in 6-well plates containing media without M-CSF or GM-CSF. The cells were treated with or without type I IFN (1,000 U/ml) for 8 h or IFN-γ (20 ng/ml) and LPS (100 ng/ml) for 24 h. Viperin expression in BMDMs was detected by immunoblot using monoclonal antibody specific to viperin (MaP.VIP). β-actin served as a protein-loading control.BM, bone marrow cells; BM+M-CSF, BMDMs differentiated with M-CSF; BM+GM-CSF, BMDMs differentiated with GM-CSF under the indicated conditions.
|
Viperin has little effect on the phenotypes of BMDMs.
To examine the effects of viperin on expression of macrophage markers and inflammatory cytokines in BMDMs, we isolated bone marrow cells from WT and viperin KO mice and differentiated into BMDMs in GM-CSF or M-CSF conditioned media. There were little differences in the expression levels of iNOS, Cd80, and Cd86 known as M1 macrophage markers, Tnf-a, Il-1b, Il-6, Il-12, and Il-23 known as M1 macrophage cytokines, Cd163, Cd206, and Arg-1 known as M2 macrophage markers, and Il-10 and Tgf-b known as M2 macrophage cytokines between WT and viperin KO BMDMs differentiated with GM-CSF (Fig. 2A). Similar results were obtained from WT and viperin KO BMDMs differentiated with M-CSF, although the expression levels of Tnf-a and Il-23 cytokines in viperin KO BMDMs had a twofold decrease (Fig. 2B). The results indicated that viperin expression is not required for the phenotypes of BMDMs differentiated with GM-CSF or M-CSF.
 | Figure 2Viperin effects on the BMDMs differentiated with M-CSF or GM-CSF. The bone marrow cells were isolated from WT and viperin KO C57BL/6 mice and differentiated into BMDMs in conditioned media. (A, B) The effects of viperin on expression of macrophage markers and cytokines. The mRNA expression levels of M1 macrophage markers and cytokines (left) and M2 macrophage markers and cytokines (right) in BMDMs differentiated with GM-CSF (A) or M-CSF (B) were measured by quantitative RT-PCR and normalized to β-actin mRNA. Data are presented as mean±SEM of triplicate samples and are representative of two individual experiments.
*p<0.05.
|
Viperin deficiency promotes gene expression of macrophage markers and cytokines in the polarized M1 and M2 macrophages.
It is well known that IFN-γ, LPS, and IL-4 stimulation can differentially induce complete M1 and M2 polarization (23). To examine viperin effects in completely polarized M1 and M2 macrophages, we stimulated WT and viperin KO BMDMs with IFN-γ and LPS for 24 h and with IL-4 for 48 h. Viperin expression was highly increased in M1 macrophages, indicating that viperin, an IFN-inducible protein is expressed by Toll-like receptor and IFN signaling pathways under IFN-γ and LPS treatments. However, viperin expression was reduced in M2 macrophages, suggesting that its expression is unable to be maintained by IL-4 in the absence of M-CSF (Fig. 3A). Polarization of BMDMs into M1 and M2 macrophages was assessed by measuring the expression levels of M1 and M2 markers (Fig. 3B). As expected, iNOS, an M1 marker was upregulated in both WT and viperin KO M1 macrophages but not in M2 macrophages. Arg-1, an M2 marker was upregulated in both WT and viperin KO M2 macrophages but not in M1 macrophages as well. Interestingly, the expression levels of iNOS and Arg-1 in viperin KO M1 and M2 macrophages were highly increased, when compared to those in WT macrophages (Fig. 3B). To further analyze this phenotype, we examined gene expression of macrophage markers and cytokines in WT and viperin KO M1 and M2 macrophages (Fig. 3C). The expression levels of M1 macrophage markers, iNOS, Cd80, and Cd86 and M1 cytokines, Tnf-a, Il-1b, Il-6, Il-12, and Il-23 in viperin KO M1 macrophages were significantly higher than those in WT M1 macrophages. Similar to these patterns, the expression levels of M2 macrophage markers, Cd163, Cd206, and Arg-1 and M2 cytokines, Il-10 and Tgf-b in viperin KO M2 macrophages were also increased when compared to those in WT M2 macrophages. The results indicated that viperin deficiency promotes the phenotypes of M1 and M2 macrophages, suggesting that viperin expression may play a key role as a regulator in macrophage polarization.
 | Figure 3Viperin effects on the polarized M1 and M2 macrophages. The bone marrow cells isolated from WT and viperin KO C57BL/6 mice were differentiated into BMDMs in M-CSF conditioned media for 7 days. The BMDMs were washed and plated in media without M-CSF. The cells were polarized to M1 macrophages by treatment with LPS (100 ng/ml) and IFN-γ (20 ng/ml) for 24 h and to M2 macrophages by treatment with IL-4 (20 ng/ml) for 48 h. (A) Viperin expression in M1 and M2 macrophages. The mRNA levels of viperin in M1 macrophages (left) and M2 macrophages (right) were measured by quantitative RT-PCR and normalized to β-actin mRNA. The mRNA level of viperin in BMDMs was used for a control. (B) Polarization of BMDMs into M1 and M2 macrophages. The polarization of WT and viperin KO macrophages was confirmed by the increased expression levels of M1 and M2 specific markers. iNOS, an M1 marker; Arg-1, an M2 marker. The mRNA levels in BMDMs were used for controls. (C) Viperin effects on the polarized macrophages. The mRNA expression levels of M1 markers and cytokines (left) and M2 markers and cytokines (right) in WT and viperin KO M1 and M2 macrophages were measured. The mRNA levels in WT M1 and M2 macrophages were used for controls. iNOS, Cd80, and Cd86, M1 markers; Tnf-a, Il-1b, Il-6, Il-12, and Il-23, M1 cytokines; Cd163, Cd206, and Arg-1, M2 markers; Il-10 and Tgf-b, M2 cytokines. (D) IRF effects on the polarized macrophages. The mRNA expression levels of IRFs in WT and viperin KO M1 and M2 macrophages were measured. The mRNA levels in WT M1 and M2 macrophages were used for controls. Irf-1, Irf-5, and Irf-8, M1-related transcriptional factors; Irf-3 and Irf-4, M2-related transcriptional factors. Data are presented as mean±SEM of triplicate samples and are representative of two individual experiments.
*p<0.05; **p<0.01.
|
It is also known that interferon regulatory factors (IRFs) have a crucial role in the polarization of M1 and M2 macrophages (232425). We examined gene expression of IRFs in WT and viperin KO M1 and M2 macrophages (Fig. 3D). The expression levels of Irf-1, Irf-5, and Irf-8 known as M1-related transcriptional factors and Irf-3 and Irf-4 known as M2-related transcriptional factors in WT M1 and M2 macrophages were similar to those of viperin KO macrophages. The results indicated that IRFs are upstream molecules of viperin in the IFN signaling pathway.
Viperin deficiency enhances macrophage marker protein expression and cytokine secretion in the polarized M1 and M2 macrophages.
To confirm the effect of viperin expression on macrophage polarization, we investigated M1 and M2 marker protein expression on cell surface and cytokine secretion using flow cytometry. The expression levels of M1 marker proteins, MHC class II and CD86 and an M2 marker protein, CD206 in viperin KO M1 and M2 macrophages were increased, when compared to those in WT macrophages (Fig. 4A). Furthermore, in cytometric bead array, the secretion levels of M1 cytokines, TNF-α and IL-6 and an M2 cytokine, IL-10 in viperin KO M1 and M2 macrophages were higher than those in WT macrophages (Fig. 4B). The data indicated that viperin expression indeed regulates expression of macrophage markers and secretion of cytokines in the polarized M1 and M2 macrophages.
 | Figure 4Viperin deficiency enhances expression of macrophage marker proteins and secretion of cytokines in the polarized macrophages. (A) The effect of viperin on expression of M1 and M2 marker proteins on cell surface. WT and viperin KO BMDMs were polarized into M1 and M2 macrophages as described above. The cells were stained with fluorescently-conjugated antibodies specific to MHC II, CD86, and CD206 and analyzed by flow cytometry. Left cytograms represent staining with an isotype-matched control antibody of irrelevant specificity. Right fraction on each cytogram indicates cell populations expressing M1 and M2 specific markers. MHC II and CD86, M1 markers; CD206, an M2 marker. (B) The effect of viperin on secretion of M1 and M2 cytokines. Cytometric bead array was performed. Open bars indicate cytokines secreted from WT M1 and M2 macrophages and closed bars represent cytokines secreted from viperin KO M1 and M2 macrophages. TNF-α and IL-6, M1 cytokines; IL-10, an M2 cytokine. Data are presented as concentration of cytokines secreted from M1 and M2 macrophages and are representative of 2 individual experiments.FSC-H, forward scatter height.
|
DISCUSSION
Viperin is highly induced in primary human and mouse macrophages treated with IFN-γ and in mouse macrophages of lymphoid organs acutely infected with lymphocytic choriomeningitis virus (81326). Our study showed that viperin is not only expressed in murine bone marrow cells, but also expressed in BMDMs differentiated with M-CSF in the absence of IFN treatment or virus infection. However, the basal expression of viperin did not affect the phenotypes, at least expression of macrophage markers and cytokines, between WT and viperin KO BMDMs. It suggested that: 1) the viperin expression level is not enough to alter the phenotypes of BMDMs, 2) viperin requires expression of other factors to affect the phenotypes of BMDMs, and 3) viperin expression might be involved in other functions including metabolic regulation rather than in immunological phenotypes of BMDMs. Furthermore, we demonstrated that viperin deficiency facilitates expression of macrophage markers and secretion of cytokines in M1 and M2 macrophages, implying a regulatory role of viperin in macrophage differentiation and polarization. The data suggested that viperin expression determines immunological functions of the completely differentiated macrophages, M1 and M2 macrophages which are activated with stimuli such as IFNs and virus-induced inflammatory cytokines in certain environment.
The main functions of macrophages are phagocytosis, endocytosis, secretion, and microbial killing (27). M1 and M2 macrophages have enhanced functional capacity compared with undifferentiated macrophages. Viperin effects on specific functions of M1 and M2 macrophages such as phagocytosis and endocytosis need to be further elucidated.
The role of viperin in M1 and M2 macrophages can be considered in macrophage-associated diseases such as asthma and atherosclerosis. When macrophages are exposed to local microenvironments, they are recruited to the sites and polarized into M1 or M2 phenotypes. Polarization of macrophages has been shown to have a crucial impact on asthma pathogenesis (28). Recent studies also showed that M2 macrophages play a central role in allergic asthma through their bidirectional interactions with immune cells and inflammatory mediators (29). In addition, it is well known that M1 macrophages are more likely to lead to an acute atherothrombotic vascular event (30), while M2 macrophages are associated with tissue repair and phagocytic capacities, enhancing plaque stability (31). A recent study showed that Irgm1 promotes murine M1 but not M2 macrophage polarization and its haplodeficiency renders ApoE KO mice resistant to atherosclerosis (32). Therefore, our data suggested that viperin regulation in macrophage polarization provides a potentially effective therapeutic strategy for macrophage-associated diseases.
Given the opposite roles of M1 and M2 macrophages in inflammatory responses and the effects of viperin on both of these polarized macrophages, viperin expression may act differently for the whole body in terms of protection or pathogenesis. Viperin expression may play a prophylactic role for inflammatory pathogenesis, including macrophage-associated diseases, by reducing the signature gene expression and cytokine production of M1 macrophages, although its expression can reduce the protective role of M1 macrophages in infections. On the other hand, viperin expression in M2 macrophages may function inversely.
In conclusion, we demonstrated that viperin participates in the polarization of macrophages. Viperin deficiency could increase the expression of M1 and M2 macrophage markers and cytokines. Our findings suggest that viperin may play a critical role in differentiation of macrophages and determination of phenotypes of M1 and M2 macrophages and provide clues to macrophage-related diseases.
 XML Download
XML Download