

 PDF
PDF Citation
Citation Print
Print


Abbreviations
CI
confidence interval
EOC
epithelial ovarian cancer
GEO
Gene Expression Omnibus
HGSOC
high-grade serous ovarian cancer
HR
hazard ratio
LGSOC
low-grade serous ovarian cancer
NCBI
National Center for Biotechnology Information
OC
ovarian cancer
OS
overall survival
PFS
progression-free survival
RFS
recurrence-free survival
TCGA
The Cancer Genome Atlas
TP53
tumor suppressor protein p53
TP53m
tumor suppressor protein p53 mutant
TP53WT
tumor suppressor protein p53 wild-type
INTRODUCTION
Ovarian cancer (OC) is the fifth leading cause of cancer deaths among women with 22,240 new cases and 14,070 deaths estimated in the US in 2018 (1). OC alone accounts for 5% of cancer deaths, which is due to the fact that early-stage OC is asymptomatic, thus when diagnosed it has frequently already spread throughout the abdominal cavity (2). Although the 5-year survival rate of OC is 92% for women diagnosed at an early-localized stage, it has low survival rate, only 17% to 28% for those with advanced-diseased stage (3). OC has been classified based on cell of origin, such as epithelial ovarian cancer (EOC) which arises in the epithelium and accounts for up to 90% of cases, and non-EOCs from germ cells for 3% and sex cord-stromal cells for 2% (4). Furthermore, EOC is histologically subdivided into 4 subtypes such as serous (up to 70%), endometrioid (10%), mucinous (6%), and clear cell (6%) (45) that differ in their epidemiologic, genetic changes, tumor markers, and response to therapy (6). In particular, high-grade serous ovarian cancer (HGSOC) is highly-aggressive, is diagnosed at advanced-stage, and has a poor prognosis (7). A frequent molecular alteration that accounts to over 95% of cases of HGSOC is the mutation in the tumor suppressor protein p53 (TP53) (89). In contrast, low-grade serous ovarian cancer (LGSOC) as well as other subtypes, such as mucinous, endometrioid and clear cells frequently do not carry TP53 mutations (1011).
To increase the survival rate of advanced OC, we need to identify molecular drivers that can serve as prognostic markers. Chemokines and their receptors have recently been receiving attention due to their possible involvement in OC progression and metastasis (1213). Chemokines, chemotactic cytokines interacting with G-protein-coupled receptors, contribute to cell proliferation, inflammation, metastasis, and tumorigenesis (141516). TP53 mutations play a significant role in shifting the effects of inflammation toward oncogenic outcomes, making cancer cells more aggressive in response to inflammatory cytokines (17). We have shown that the frequent mutation of TP53 in OC enhances proinflammatory chemokines, leading to inflammatory tumor microenvironment (18). In addition, TP53 inactivation during carcinogenesis affects immune surveillance by interfering with chemokine expression (19).
To date, there have been no prior reports on patient survival based on TP53 status and chemokines in OC. Here, we investigated the chemokine network in different subtypes of human EOC, focusing on whether there is association in TP53 status and chemokine network and how this correlates with OC survival.
MATERIALS AND METHODS
Data analysis
Data analysis was performed on publicly available microarray data-sets that were deposited in the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) (http://www.ncbi.nlm.nih.gov/geo/) database under accession numbers GSE6008 and GSE63885. For GSE6008, RNA expression data were from 99 individual OC cases (37 endometrioid, 41 serous, 13 mucinous, and 8 clear cell carcinomas) and 4 individual normal ovarian samples. GSE63885 is gene expression data from 101 OC surgical samples, including 71 serous carcinomas, for which somatic TP53 gene mutation status was available.
We utilized Gitools 2.3.1 (http://www.gitools.org) based on Oracle Java 7, an open-source tool to perform Genomic Data Analysis and Visualization as interactive heat-maps (20). Kaplan-Meier plotter database (http://kmplot.com/analysis/index.php?p=service&cancer=ovar) was utilized to evaluate progression-free survival (PFS) and overall survival (OS) using proportional hazards regression to estimate hazard ratios (HRs) and 95% confidence intervals (CIs) based on gene expression profile of chemokines among 1,656 OC patients from GEO and The Cancer Genome Atlas (TCGA); chemokines were specified with probe sets (Affymetrix HG-U133A, HG-U133A 2.0, and HG-U133 Plus 2.0 microarrays) (21). TP53 alteration frequency profile across many types of cancers was acquired from cBioPortal (www.cbioportal.org) (2223); studies with <25% of altered samples were not included. Data on expression levels were analyzed using Student's t-test and 1-way ANOVA as appropriate. If statistical significance (p≤0.05) was indicated by ANOVA, then data were further analyzed using Tukey's pairwise comparisons to detect specific group differences.
RESULTS AND DISCUSSION
TP53 is the most frequently altered gene in many types of malignancies, with mutations in at least 50% of human cancers. First, we checked the alteration frequency in TP53 across different types of cancers. TP53 mutation was common across many malignancies, while deletion, amplification and multiple alterations in TP53 occurred in only a few types of cancers (Fig. 1). OC in the published TCGA dataset, which contains only HGSOC (9), had the highest percentage of TP53 mutation (Fig. 1). Even all of de novo HGSOC are supposed to contain TP53 somatic mutations or deletions, except for the rare HGSOC that develop from a low-grade serous tumor precursor (24).
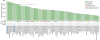 | Figure 1Analysis of alteration frequency (%) in TP53 across many cancer types. The 150 studies deposited in cBioPortal (www.cbioportal.org) were assessed to analyze the TP53% alteration frequency profile. The 68 studies with <25% alteration frequency were filtered out to create this graph. Green, blue, red and gray bars indicate mutation, deletion, amplification and multiple alterations in TP53, respectively. The dot line indicates 80%.
|
OC is associated with chronic inflammation (25). Previously, we reported that loss or mutation of TP53 may promote tumor progression by enhancing proinflammatory chemokines in OC (18). Chemokines and their receptors are differentially expressed in tumors, influencing cancer progression. We then evaluated whether the different subtypes of EOC would show different patterns of chemokine expression profiles based on publicly available GEO data (GSE6008). Clear cell OC has high CXCL14 expression, while mucinous OC had high CCL15 and CCL20 expression (Fig. 2). Functional roles of these chemokines in clear cell and mucinous OC are largely unknown. On the other hand, normal ovarian samples highly expressed CCL18 and CXCL12 compared to other EOC subtypes (Fig. 2). Consistently, normal ovarian stroma expressed a higher level of CXCL12 compared to EOC (26). Although serum CCL18 was elevated in patients with EOC (27), patients with higher CCL18 in tumor samples had better OS in OC (HR=0.88; 95% CI=0.77–1.00; n=1,656) based on our analysis. Data of TP53 mutation in clear cell and mucinous OC subtypes are not available for statistical analysis because of limited data. Next, we classified endometrioid EOC into TP53 wild-type (TP53WT) and TP53 mutant (TP53m) and assessed chemokine signatures. CXCL14 was highly expressed in TP53WT endometrioid OC compared to TP53m (Fig. 3A). Because of limited data to identify TP53WT and TP53m endometrioid OC patients for OS measurement, we used total endometrioid OC to investigate OS and PFS based on the expression levels of CXCL14. Patients with high expression of CXCL14 has better PFS (HR=0.22; 95% CI=0.08–0.63) but unchanged OS in endometrioid OC (Fig. 3B). CXCL14 protein level is positively correlated to OS of breast cancer patients (28). Moreover, CXCL14 acts as a tumor suppressor gene that is epigenetically silenced during lung tumorigenesis, and the re-expression of CXCL14 leads to increased cell death and reduced growth of lung tumor xenografts (29). CXCL14 is a potent angiostatic chemokine which prevents chemokine- and growth factor-induced angiogenesis (30). High expression levels of CXCL14 in clear cell and endometrioid OC may be one reason why these subtypes have better prognosis than serous OC. Interestingly, CXCL14 was reported to be expressed in normal tissue, such as the epithelia (31), and OC stroma (26).
 | Figure 2Chemokine and chemokine receptor signatures in EOC subtypes. Heatmap of chemokine expression profiles in EOC subtypes, including clear cell (n=8, yellow), endometrioid (n=37, blue), mucinous (n=13, green), and serous (n=41, red), and normal ovarian samples (n=4, black) from the NCBI GEO (http://www.ncbi.nlm.nih.gov/geo/) database (GSE6008) using Gitools 2.3.1. Bold yellow, green and black letters indicate dominant chemokines in clear cell, mucinous and normal samples, respectively. The right panel indicates statistical analysis of chemokine expression intensities using ANOVA and Tukey's pairwise comparisons.
|
 | Figure 3Chemokine and chemokine receptor signatures in TP53WT and TP53m endometrioid OC. (A) Heatmap of chemokine and chemokine receptor expression profiles in TP53WT (n=22) and TP53m (n=15) endometrioid OC from NCBI GEO (http://www.ncbi.nlm.nih.gov/geo/) database (GSE6008) using Gitools 2.3.1. Blue letter indicates the dominant chemokine in TP53WT endometrioid OC. Right panel indicates statistical analysis of chemokine expression intensities using Student's t-test. (B) Kaplan-Meier plots for OS and PFS based on expression levels of CXCL14 in endometrioid OC patients from GEO and TCGA (probes; Affymetrix HG-U133A, HG-U133A 2.0, and HG-U133 Plus 2.0 microarrays). Black and red letters indicate low and high expression of CXCL14, respectively.
|
We determined the chemokine signature in TP53WT and TP53m serous EOC subtype from GEO dataset (GSE63885). Chemokines, such as CCL8, CCL20, CXCL10 and CXCL11, are highly expressed in TP53m compared to TP53WT serous OC (Fig. 4). Our previous study showed that CCL20 is expressed dominantly in CXCR2-driven OC, leading to shorter survival, greater tumor spread in the peritoneal cavity, and larger tumor weights in xenograft OC models (26). Chemokines can exert pro- and anti-tumorigenic effects, depending upon the context (32). CXCL10 is thought to impair angiogenesis and has anti-tumor actions (3334). Increased expression of CXCL10 had also been linked with advanced human malignancies such as melanoma (35) and OC (36). The chemokine network is involved in the metastasis of TP53WT and TP53m malignancies. The mutant TP53 protein enhances the secretion of CXC chemokines via the NF‐κB pathway, leading to increase cell migration (37). Particularly, CXCL1 and CXCL8, implicated in cancer invasiveness and angiogenesis, are downregulated in TP53WT-transfected OC (18). In addition, increased expression levels of CXCL5, CXCL8, and CXCL12 were found in TP53m cells compared to TP53WT cells (37). In several malignancies such as lung, melanoma and breast cancer, knockdown of the mutant TP53 protein led to reduced chemokine levels and cell migration (37).
 | Figure 4Chemokine and chemokine receptor signatures in TP53WT and TP53m serous OC. (A) Heatmap of chemokine and chemokine receptor expression profiles in TP53WT (n=9) and TP53m (n=62) serous EOC from NCBI GEO (http://www.ncbi.nlm.nih.gov/geo/) database (GSE63885) using Gitools 2.3.1. Red letters indicate dominant chemokines in TP53m serous OC. Right panel indicates statistical analysis of chemokine expression intensities using Student's t-test.
|
Finally, we utilized datasets available in the Kaplan-Meier plotter database to evaluate OS based on chemokine signatures in TP53WT and TP53m serous OC. Tables 1 and 2 show HRs for OS based on chemokine and chemokine receptor signatures in TP53WT and TP53m serous OC, respectively. High expression levels of CXCL12 (HR=1.84; 95% CI=1.05–3.22) and CCL21 (HR=1.76; 95% CI=1.01–3.06) were associated with poor OS in TP53WT serous OC (Fig. 5A and B). In a similar study, EOC patients with highly expressed CXCL12 had poor survival compared to patients with lower levels (13). High expression of CXCR2 (HR=1.34; 95% CI=1.06–1.69) was associated with poor OS in TP53m serous OC, while high expressions of CXCL9 (HR=0.78; 95% CI=0.62–0.98), CCL5 (HR=0.77; 95% CI=0.61–0.98), CXCR4 (HR=0.75; 95% CI=0.59–0.94), CXCL11 (HR=0.72; 95% CI=0.57–0.91), and CXCL13 (HR=0.64; 95% CI=0.51–0.81) were associated with better OS (Fig. 5A and C). CXCR2 was correlated with poor OS and recurrence-free survival (RFS) in non-metastatic patients with non-clear cell renal carcinoma patients (38). In addition, analysis of 12 studies with a total of 2,461 cancer patients including laryngeal squamous cell carcinoma, lung cancer and gastric cancer indicated that high expression level of CXCR2 was associated with poor OS and RFS (39). Overexpression of CXCR2 was associated with poor OS and disease-free survival of patients with HGSOC (40). These studies suggest poorer OS in TP53m serous OC with high levels of CXCR2 (Table 2, Fig. 5A and C). CXCL9 and CCL5 released by IL18-stimulated natural killer cells aid in the recruitment of effector T cells by promoting type I response against cancer (414243). Consistently, high expression levels of CXCL9 and CCL5 in TP53m have better OS (Table 1, Fig. 5A and C). EOC patients with high expression of CXCL12 and low expression of CXCR4 showed worse survival (13). Accordingly, high expression of CXCL12 in TP53WT serous OC has worse OS, while high levels of CXCR4 in TP53m serous OC has better OS (Tables 1 and 2, Fig. 5).
Table 1
HRs for OS based on expression levels of chemokines in serous OC
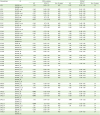
![]()
Table 2
HRs for OS based on expression levels chemokine receptors in serous OC

![]()
 | Figure 5Chemokine and chemokine receptor signatures associated with OS in TP53WT and TP53m serous OC. (A) HR of chemokines and chemokine receptors that were either positively or negatively related with OS in TP53WT and TP53m serous OC using GEO and TCGA datasets available in the Kaplan-Meier plotter database (http://kmplot.com/analysis/index.php?p=service&cancer=ovar). (B) Kaplan-Meier OS for CXCL12 and CCL21 in TP53WT serous OC (n=91). (C) Kaplan-Meier OS for CXCR2, CXCL9, CCL5, CXCR4, CXCL11 and CXCL13 in TP53m serous OC (n=493). Blue and red letters indicate low and high expression of chemokines and chemokine receptors, respectively.
|
The present study has several limitations. Less common OC subtypes such as clear cell, mucinous and endometrioid OC had limited data available. Also, we could not perform OS analysis for TP53WT and TP53m endometrioid OC because of limited data number. Even OS analysis for serous OC, a representative OC, has unbalanced number between TP53WT (n=91) and TP53m (n=493) serous OC because of more frequent mutation of OC compared to other cancer types (Fig. 1). We cannot exclude the possibility that these TP53WT serous OC most likely represent LGSOC (24), since published TCGA data encompassing only HGSOC show that above 95% of tumors are TP53m (9). Therefore, differences in chemokine signatures between, and survival of patients with TP53WT and TP53m serous OC may be a reflection of different subtype. Further studies and additional data will help to overcome these limitations.
In conclusion, a distinct chemokine signature may be seen in TP53WT and TP53m OC and be associated with better or worse OS, suggesting possible utility as a biomarker for OC prognosis. In particular, CXCR2 has worse OS in TP53m serous OC, the most representative OC, and may be a potential target for OC therapy.
 XML Download
XML Download