

 PDF
PDF Citation
Citation Print
Print


Abbreviations
ARE
antioxidant response element
ATP
adenosine-5′-triphosphate
DAMP
damage-associated molecular pattern
DC
dendritic cell
DT
diphtheria toxin
HMGB1
high mobility group box 1
HO
heme oxygenase
ICAM
intercellular adhesion molecule
IFN
interferon
IRI
ischemia-reperfusion injury
KC
Kupffer cell
Keap1
Kelch-like ECH-associated protein 1
LPS
lipopolysaccharide
LT
liver transplantation
NF
nuclear factor
NK
natural killer
NKT
natural killer T
NLRP3
nucleotide-binding domain leucine-rich repeat containing family pyrin domain containing 3
Nrf2
nuclear factor erythroid 2-related factor 2
PRR
pattern recognition receptor
ROS
reactive oxygen species
TLR
Toll-like receptor
TIM
T cell, immunoglobulin, mucin-containing molecules
TNF
tumor necrosis factor
WT
wild type
INTRODUCTION
With improvements in surgical procedures, immunosuppression techniques and perioperative care, liver transplantation (LT) has evolved into a definitive treatment for all types of end-stage liver diseases and is performed in over 80 countries (1). Although the 1-year survival rates of patients undergoing LT have exceeded 80% (1), some issues are still troublesome limitations, including ischemia-reperfusion injury (IRI). IRI, occurring during hemorrhagic shock, liver resection and LT, is a biphasic pathophysiological process that includes the initial ischemic liver damage and subsequent reperfusion injury. The ischemia insult characterized with the restriction of the blood supply during organ procurement can cause the cellular metabolic stress due to the hypoxia, adenosine-5′-triphosphate (ATP) and glycogen consumption, mitochondrial dysfunction, which all contributes to the initial cell deaths. The reperfusion injury, on the other hand, refers to the phenomenon that after the restoration of the blood flow and reoxygenation, the livers usually suffer from a severer damage, which have a great relationship with not only the metabolic disturbance and oxygen radicals, but also the inflammation reaction cascade involving with various immune cells, complements and cytokines or chemokines they produced. Researches have revealed that many pathological processes are involved in the development of IRI, including the increasing vascular permeability and leakage, activation of cell death programs (apoptosis, autophagy-associated cell death, and necrosis), transcriptional reprogramming, generation of reactive oxygen species (ROS) and nitrogen species, and the activation of the innate and adaptive immune network (234). It is of particular note that the hepatocellular damage concerning IRI is crucial to the occurrence of poor early grafts function, even primary nonfunction. Furthermore, IRI may also be responsible for a considerable portion of postoperative acute and chronic rejection. Therefore, grafts with longer ischemia time are usually discarded, which further worsen the organ shortage (24). Unfortunately, the mechanisms of IRI remain much unknown and no effective therapy is available in the clinical application despite of many great advancements in basic animal models' researches.
The liver is a unique organ that harbors various types of innate cells, including resident macrophages (Kupffer cells; KCs), dendritic cells (DCs), natural killer (NK) cells, and natural killer T (NKT) cells (56), which all play important roles in IRI. In this review, we aimed to summarize the current understanding of KCs that are the predominant players in sterile inflammation and highlighted the activation and roles of these cells in IRI.
QUIESCENT STATE OF KCs
Proper macrophage nomenclature is intensively debated at present, but basically the hepatic macrophages origin from three sources (Fig. 1) (789). KCs are a group of hepatic macrophages and they are the largest population of tissue-resident macrophages in the body. They reside within the liver sinusoid and play a pivotal role in the immune response. In the steady state condition, the intimate contact with circulation blood flowing from the portal vein or hepatic artery allows them to phagocytize majority of the pathogens such as microorganisms and their products, as well as those deriving from the gastrointestinal ducts. Besides, they are also in charge of the clearance of the other substance including aged erythrocytes, cell debris, tumor cells, immune complexes (10111213). It is now convinced that the KCs in a healthy liver exhibit the ‘tolerogenic’ phenotypes, which maintain the immunological tolerance. However, in the diseases state, they may undergo phenotype switch to participant in the immune reaction. Basically, pro-inflammatory cytokines such as tumor necrosis factor (TNF)-α, IL-6, IL-12, IL-1β, and inducible nitric oxide (NO) synthase are secreted by a group of KCs while another type may work contrarily to increase the expression of anti-inflammatory mediators. Furthermore, they could also interact with other immune cells to facility the response (1213). Lots of works has been done to elaborate the contributions of KCs to the liver diseases, such as fibrosis (12), alcoholic and nonalcoholic fatty liver diseases (13), hepatotropic virus infection (14), and cholestatic liver injury (15).
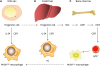 | Figure 1Origins of hepatic macrophages.The hepatic macrophages in adults derives from at least 3 sources. (A) Yolk sac. It can produce progenitors that populate all tissues and that have progeny that persist throughout life as F4/80 high resident macrophages. These lineages are mainly regulated by CSF1R and its ligands, IL-34 and CSF1. (B) Fetal liver. This is less well defined but seems to contribute to the production of adult KCs. (C) Bone marrow. The source which give rise to circulating monocytes and progeny F4/80 low macrophages. Macrophages that are F4/80 low emanate from Ly6c1 monocytes. The exact role of the patrolling Ly6c negative macrophages, and the contribution of fetal liver to adult tissue macrophages remains unclear.
CSF1, colony-stimulating factor 1; HSC, hepatic stellate cell; GMP, granulocyte-monocyte progenitor; MDP, monocyte dendritic cell progenitor.
|
In conclusion, in steady state condition, the KCs exhibits the ‘tolerogenic’ type and are in charge of the clearance of pathogens. In the diseases, they could change phenotype and play a role in the immune reaction.
ACTIVATION OF KCs
Until now, 2 different stages of IRI have been identified (24). In the ischemia injury stage, hypoxia and accompanying metabolic disturbances induce initial cell death, which releases diverse ‘alarmins’ such as damage-associated molecular patterns (DAMPs), ROS, and DNA fragments. In the reperfusion injury stage, in response to stimulation by alarmins, resident immune cells are activated, and these cells facilitate the generation of a pro-inflammatory milieu, which in turn amplifies and sustains immune cascades by recruiting immune cells from the circulation.
Toll-like receptors (TLRs) are a family of pattern recognition receptors (PRRs) that are expressed in various liver cells and can trigger immune inflammation in response to DAMPs and other alarmins (616). KCs express TLR2, TLR3, TLR4 and TLR9 on the cell surface or in endosomes/lysosomes (17). TLR4 signaling is a well-studied pathway that is critical not only for nonparenchymal cells, which are involved in the initiation of innate and adaptive immune responses (18), but also for parenchymal cells, which release danger signals such as high mobility group box 1 (HMGB1) (1920).
HMGB1, the key endogenous DAMP molecule in liver immune activation, is upregulated as early as 1h after reperfusion (21). As a nuclear factor (NF), HMGB1 is actively released by hypoxic hepatocytes during IRI, and the process is promoted by TLR4-dependent ROS production and downstream calcium/calmodulin-dependent protein kinases-mediated signaling (22). With the binding of TLR4 and its major ligand HMGB1, KCs are activated. Although the inhibition of the activity and release of HMGB1 exerts a protective effect during IRI (2123), the biological role of HMGB1 seems more complex, as hepatocyte-specific HMGB1 deletion increases nuclear instability and histone release and, consequently, worsens liver IRI through the activation of TLR9 (24). Hence, during liver IRI, HMGB1 may have 2 roles, namely, a beneficial intracellular role and an injurious extracellular role (25).
Innate immune activation involves a complex network consisting of multiple cells and molecules (26). In addition to HMGB1, other DAMPs including histones, DNA fragments and ATP, mitochondrial ROS and complement proteins stimulate KCs and other distinctive immune cells via different PRRs, such as TLR9 (recognizing histones) (26), TLR3(recognizing RNA) (2728), TLR4 (recognizing heat shock protein [HSP]-70) (29), and nucleotide-binding domain leucine-rich repeat containing family pyrin domain containing 3 (NLRP3) (recognizing ATP) (303132).
Lipopolysaccharide (LPS) is a gut-derived endotoxin that can translocate into the hepatic circulation during IRI because the occlusion of portal vein blood flow increases the permeability of the intestinal wall. LPS has been reported to bind to LPS-binding proteins, engage CD14 and activate KCs through the mediation of TLR4 signaling (33). However, whether LPS can trigger or sustain the activation of KCs remains to be determined because an increase in LPS was detected in late phases of IRI and because LPS-neutralizing agents failed to protect mouse liver during early phases of IRI (34). It is thus likely that this gut-derived endotoxin sustains immune inflammation (2).
In conclusion, KCs are activated by the DAMPs or other alarmins through PRRs.
DUAL EFFECT OF ACTIVATED KCs IN INNATE AND ADAPTIVE IMMUNE SIGNALING
The response of activated KCs to reperfusion is bifunctional, with the secretion of both pro-inflammatory and anti-inflammatory factors (Fig. 2). TNF-α, produced by KCs, is one of the principal cytokines in IRI. TNF-α exacerbates damage and induces apoptosis by directly binding to the TNF receptor on the surface of hepatocytes and activating the NF-ĸB and c-Jun N-terminal kinase (JNK) pathways (35). A recent study also revealed that TNF-α-induced tissue injury can be mediated by the peroxisome proliferator-activated receptor-γ co-activator (PGC)-1α/mitofusion (Mfn)-2 pathway, which regulates mitochondrial functions and energy metabolism (36). Furthermore, TNF-α can increase the severity of inflammation by upregulating the expression of other inflammatory factors including ROS, intercellular adhesion molecule (ICAM)-1, vascular adhesion molecule (VCAM)-1 and P-selectin and inducing the recruitment of neutrophils (3738). Notably, KCs and TNF-α also participate in the mediation of hepatic injury after remote organ IRI (39). TNF-α suppression (40) or gene silencing (41) play marked protective roles in animal IRI models; however, improvements have also been observed with TNF-α preconditioning (42), indicating the dual roles played by this molecule (43).
 | Figure 2Dual effect of activated KCs in IRI.The response of activated KCs to reperfusion is bifunctional with the secretion of both pro-inflammatory and anti-inflammatory factors. TNF-α, IL-1β, IL-18, IFN-γ, and IL-12 secreted by KCs show a pro-inflammatory effect, in contrast, IL-10 and HO-1/Nfr2 pathway contribute to the anti-inflammatory regulation.
JNK, c-Jun N-terminal kinase; PGC-1α, peroxisome proliferator-activated receptor-γ co-activator 1α; Mfn-2, mitofusion 2; VCAM-1, vascular adhesion molecule 1; MIP-2, macrophage inflammatory protein 2.
|
Other quintessential pro-inflammatory cytokines include IL-1β and IL-18, whose synthesis, activation and release are controlled by caspase-1 (44). NLRP3 is another PRR that is mainly expressed on KCs. NLRP3 can be activated in response to stimulation by DAMPs, and it induces the assembly of the NLPR3 inflammasome with apoptosis-associated speck-like protein containing a caspase activation and recruitment domain (ASC) and pro-caspase1. Inflammasomes are multiprotein complexes that can recognize damage signals and directly activate caspase-1 (324445). Studies have demonstrated that endogenous extracellular histones binding to TLR9 can activate the NLRP3 inflammasome, which can then initiate the activation of caspase-1 and the secretion of IL-1β and IL-18 (46). This promotes the production of HMGB1 (47) and the recruitment of neutrophils and leukocytes from the circulation (3746). In addition, the absent in melanoma (AIM)-2 inflammasome, activated by double-stranded DNA in KCs, also contributes to caspase-1 activation and propagates the injury (48).
The other pro-inflammatory factors that are completely or partly derived from KCs include ROS, interferon (IFN)-γ and IL-12, which recruit immune cells, cause damage to hepatocytes or sinusoidal endothelial cells and increase the expression of other inflammatory factors and adhesion molecules (3738).
Although the inhibition of KCs has shown promising outcomes in ameliorating liver IRI in some studies (495051), KC-depleted mice have also shown an increased level of injury followed with higher mortality (5253). IL-10 is one of the primary anti-inflammatory cytokines produced by KCs, and it can suppress NF-κB activation and dramatically inhibit the expression of pro-inflammatory factors such as TNF-α, IL-1β, IFN-γ, IL-2, E-selectin, macrophage inflammatory protein (MIP)-2, and ICAM-1 (5253545556). Moreover, TLR4 suppression and apoptosis inhibition are also observed in animals subjected to adenovirus (Ad)-based viral IL-10 gene transfer (56). Furthermore, IL-10 can reduce inflammation-associated injury by upregulating the expression of heme oxygenase (HO)-1 (5657), which plays a profound cytoprotective role in liver IRI. HO-1 is predominantly secreted by KCs and can drive the differentiation of these cells, leading to an increase in the number of ‘anti-inflammatory’ macrophage populations (58). Both warm ischemia-reperfusion (IR) and orthotopic LT animal models have validated that Ad-based HO-1-transduced macrophages can attenuate liver IRI and improve survival by not only decreasing local neutrophil accumulation and pro-inflammatory factor expression but also increasing the level of IL-10. Cell apoptosis is also ameliorated under these conditions (5960).
It is well documented that the Kelch-like ECH-associated protein 1 (Keap1)/nuclear factor erythroid 2-related factor 2 (Nrf2)/antioxidant response element (ARE) pathway is responsible for the regulation of HO-1. Under normal conditions, Nrf2 is sequestered in the cytoplasm by Keap1, which facilitates its degradation via the ubiquitin-proteasome pathway. However, under stressful circumstances, in response to specific chemical signals, Nrf2 can dissociate from Keap1 and translocate into the nucleus. The binding of Nrf2 to AREs upregulates the transcription of numerous antioxidants and anti-apoptotic proteins including HO-1 (6162). Interestingly, another study revealed that nuclear HO-1 can interact with Nrf2 to enhance the stabilization of Nrf2 and, consequently, form a positive loop (63). Upstream factors such as protein kinase C (PKC) and activating transcription factor (ATF)-3 are also found to mediate the Nrf2/HO-1 pathway, and this could be an innovative strategy to manage IRI (6465). Moreover, autophagy has been shown to be involved in the protection of liver from IRI (6667) via the Nrf2/HO-1 pathway (6869).
Though the innate immune response takes the dominant role in hepatic IRI, robust adaptive immune reaction is also indispensable, which mainly depends on the CD4+ T cells (2). In the absence of exogenous antigen stimulation, CD4+ T cells can be activated by the pro-inflammatory cytokine or chemokines produced by the KCs, neutrophils and hepatocytes and recruited to the post-ischemic livers (37). What is more, the accumulation of CD4+ T cells comes rapidly after the IR and facilitate the accumulation of neutrophils via IL-17 (23770). On the other hand, the activated CD4+ T cell can in turn activate the innate immune cells including KCs and sustain the inflammation via the CD154/CD40 (2), TIM-1/TIM-4 (271). CD154 and TIM-1 are expressed in the CD4+ T cells and the corresponding ligands CD40 and TIM-4 are expressed in the macrophage and antigen-presenting cells. In contrast, the PD-1/B7-H1 (72) and TIM-3-galectin-9 pathway play a negatively stimulatory role in promoting IRI (73). That means, the KC and CD4+ T cell could interact with each other to regulate the IRI.
In summary, the KCs could play a dual effect in IRI and it could interact with the CD4+ T cells in the adaptive immune reaction.
DISTINCTIVE ROLES OF RESIDENT KCs AND INFILTRATING MACROPHAGES
Human macrophages can be classified into three subsets based on the expression of CD11b, CD68 and CD32. In contrast to macrophages in circulation or other organs expressing CD11b, liver-resident KCs are mainly CD68+ and/or CD32+ (11). In addition, CD32/CD68+ KCs can be depleted by clodronate liposomes, whereas the CD11b+ cells are sensitive to radiation and diphtheria toxin (DT) (611). Depletion of the circulation macrophages with the treatment of DT in the CD11b-DT receptor (DTR) mice resulted in IRI-resistance when compared with the untreated mouse, which presented a decreased alanine transaminase (ALT) level and preserved architecture and the damage could be restored by the adoptive infusion of macrophages, which demonstrated the pro-inflammatory role of the non-resident macrophages. However, when infusing the TIM-4 deficient macrophages, the restoration was failed, which highlighted the importance of TIM-4 in the hepatic IRI, which could be a potential therapeutic target (74).
Other innate immune cells also function in the hepatic IRI. Studies has demonstrated that both the resident and infiltrating types of DCs are involved in the hepatic IRI. Specifically, the blood recruited DCs may serve as pro-inflammatory cells as the adoptive infusion of wild-type DCs to the Flt3L KO (DC-deficient) or wild type (WT) mice increased IRI injury while the resident DCs may function in a contrary way as the grafts lacking resident DCs (Flt3L KO liver grafts) suffered severer injuries when transplanted into a WT mice compared with the WT livers transplanted into WT mice (75). Besides, those DCs may protect the liver from IRI by secreting IL-10 via TLR9 signals (76). Similar opposing roles has also been observed in NKT cells that type I NKT cells promoted the IRI injury and the type II suppressed it. When activated by the sulfatide, the type II NKT cells would reduce the production of IFN-γ from type I NKT cells and reduce the recruitment of myeloid cells, which could be a novel therapeutic strategy for IRI (77). IFN-γ is also secreted by the CD-39 mediated NK cells. Studies of Beldi (78) has revealed the deletion of CD39 on NK cell could abrogate the level of IFN-γ and attenuates hepatic IRI. Kimura et al. (79) also showed that NK cells diminished quickly in the allogeneic grafts in the rat model of transplantation but not in the syngeneic grafts, which demonstrated its role of alien recognition. Furthermore, depletion of mature NK cells improved the hepatic IRI.
In summary, other innate cells also play significant role in the IRI.
CONCLUSION AND FUTURE DIRECTIONS
In conclusion, IRI is a severe problem during LT, and it activates both the innate and adaptive immune system, leading to sterile inflammatory responses to damage hepatocytes. Liver-resident macrophages or KCs, activated by the interaction between alarmins and PRRs, play a pivotal role during IRI. Activated KCs can function as both a pro-inflammatory and an anti-inflammatory manner.
Studies investigating complex signaling pathways in KCs may aid in the development of effective therapeutic strategies to attenuate inflammation-associated injury. Specifically, inhibition of pro-inflammatory signaling combine with/or stimulation of anti-inflammatory regulators can efficiently ameliorate IRI. NF-κB is a main pathway activated by TNF-α and its suppression shows an obvious reduction effect of IRI (4053808182). Butyrate as a 4-carbon fatty acid help reduce liver IRI by inhibiting NF-κB (80). Similarly, down regulation effect of epigallocatechin-3-gallate and peptidyl-prolyl isomerase, Pin1 showed an attenuation of IRI (8384). HMGB 1 as a key mediator of pro-inflammatory pathway also been investigated to protect IRI (8586). Tanshinone IIA shows a potential role of prevention of IRI by down-regulation of the HGMB1-TLR-4/NF-κB pathway in KCs and activation of phosphatase and tensin homolog deleted on chromosome ten (PTEN)/phosphatidylinositol 3 kinase (PI3K)/AKT pathway (85). Glycyrrhizin is a natural anti-inflammatory and antiviral triterpene in clinical use. Ogiku et al. (86) figure out the expression of HGMB1 in serum and production in HGMB1 in KCs was reduced in the glycyrrhizin group compared with the control. Suppression of pro-inflammatory star mediator TNF-α by glutathione (40) and improvement of anti-inflammatory molecule IL-10 (53) were also verified their controversial effect in the process of IRI development. Functional HO-1 protein conjugated to a cell-penetrating peptide exert significant cytoprotective effect to hepatocytes in vitro (87). Some groups approve the positive influence by altering the M2 macrophage polarization (8889). The small molecules like ROS, NO also can be a power clinical treatment tools for the IRI patients after LT (9091). Interestingly, non-coding RNA miR-155 deficiency plays an effective role in attenuating liver IRI likely by regulating the activation and inflammatory response, as well as modifying the polarization of KCs (92).
Overall, the efforts to combine the repression of pro-inflammatory mediator with promotion of anti-inflammatory elements in KCs will further improve the prognosis of LT patient suffering from IRI. Recent findings motivate the search for novel therapeutic targets inside KCs to aid in moving personalized health care of IRI patients a step forward.
 XML Download
XML Download