

 PDF
PDF Citation
Citation Print
Print


Abbreviations
AMP
antimicrobial peptide
BCG
Bacillus Calmette-Guérin
CS
cigarette smoke
CTLA
cytotoxic T-lymphocyte-associated antigen
IFN
interferon
LTBI
latent tuberculosis infection
MAIT
mucosal associated invariant T
mprF
multiple peptide resistance factor
MTB
Mycobacterium tuberculosis
PD-1
programmed cell death protein 1
S. aureus
Staphylococcus aureus
SHS
second-hand smoke
TB
tuberculosis
TGF
transforming growth factor
TNF
tumor necrosis factor
Treg
T regulatory cell
TST
tuberculin skin test
INTRODUCTION
Cigarette smoking is increasing worldwide and by reasonable inference, second-hand smoke (SHS) exposure is likely increasing in parallel (1). SHS, also called environmental tobacco smoke, involuntary smoke, and passive smoke, is comprised mainly, by up to 85%, of “sidestream” smoke (the smoke given off by a burning cigarette or another tobacco product) and to a lesser extent (up to 15%) of “mainstream” smoke (the smoke inhaled and exhaled by a smoker). Not surprisingly, sidestream smoke and hence SHS contains >7,000 distinct chemical compounds including nicotine. Indeed, nicotine has been found in the exhaled breath, oral fluids, and blood of individuals exposed to SHS and second-hand vapors from electronic cigarettes (23). SHS exposure has been shown to cause many of the same diseases caused by direct smoking (4). In fact, sidestream smoke — the major component of SHS — may be more harmful, in the context of toxicity and tumorogenicity, than mainstream smoke (5).
Tuberculosis (TB) is an ongoing global public health crisis (6). The geographic overlap of the prevalence of cigarette smoking and the number of latent and active TB cases is striking (7). Mathematical models project that cigarette smoke (CS) exposure contributes to millions of TB cases and deaths worldwide and is a significant driver of the TB pandemic (89). SHS exposure is likely more common in TB endemic countries than in the U.S. because of greater smoking prevalences and less stringent public smoking bans in those countries. There appears to be no safe levels of SHS exposure (10). Indeed, small airway epithelial cells exposed in vivo to the lowest detectable level of CS — as that seen with SHS — displayed abnormal gene expression (11). Our goals are to review the epidemiological studies that examined the relationship between SHS exposure and TB as well as summarize the effects SHS/direct CS exposure have on immune cells that play a role in host defense and/or the pathogenesis of TB.
METHODS
We reviewed the literature — using PubMed searches — on SHS exposure and its association with various clinical forms of TB including latent tuberculosis infection (LTBI) and active TB (12). We also searched for the effects of SHS/direct CS on the functions of macrophages, neutrophils, T effector cells, CD8+ mucosal associated invariant T (MAIT) cells, B cells, and T regulatory cells (Tregs).
EPIDEMIOLOGIC STUDIES LINKING SHS EXPOSURE WITH TB INFECTION AND DISEASE
TB infection
As part of a larger analysis, we previously reported 5 studies that examined the association between LTBI and SHS exposure (1314151617). In a cross-sectional study of nearly 8,000 high school students, those with larger tuberculin skin test (TST) reactivity (mode up to 16 mm induration) were twice as likely to have 2 smoking parents than to have one or no smoking parents (16). In 2 separate studies from South Africa of up to 1,500 children, those who were contacts of smokers with active TB or who lived with 2 or more smokers with active TB were more likely to have a positive TST than contacts of non-smokers with active TB (1314). In over 7,000 Spaniards, those in contact with smokers diagnosed with active TB were more likely to have a positive TST than subjects in contact with non-smokers with TB (15). In 95 children from the Indian subcontinent who were contacts of adults with active TB, the determining risk factors for transmission of infection were younger age, severe malnutrition, absence of Bacillus Calmette-Guérin (BCG) vaccination, contact with a sputum-positive adult, and exposure to SHS (17). More recently, 2 studies reported meta-analyses on the association of SHS exposure and LTBI or active TB (1819). Patra and colleagues analyzed 18 studies and found a significant association between SHS and LTBI with a pooled relative risk (RR) of 1.64 (95% confidence interval [CI], 1.00–2.83) but observed great heterogeneity in the studies (18). Dogar and co-workers (19) performed a meta-analysis on 12 studies and found that SHS exposure was associated with LTBI although this did not reach statistical significance (RR, 1.19; 95% CI, 0.90–1.57); marked variability in the results were also found, which was attributed to differences in the diagnostic criteria used among the studies analyzed.
Primary progressive TB
In infants and young children, active TB cases are almost always due to progression of primary infection, since they are unlikely old enough to have LTBI (20). Four studies in children found a significant association between SHS and primary progressive TB (21222324). Altet et al. (21) also showed a greater association between primary progressive TB in children and the amount smoked in the household. In 1 study from Thailand and 2 from India, children with active TB were more likely to have had SHS exposure (222324). Patra et al. (18) performed a meta-analysis of the studies that examined the association between SHS and active TB in children and found a 3-fold greater association than those without SHS exposure.
Active TB in adults
We previously summarized the numerous studies that investigated the association between cigarette smoking and active TB with the vast majority noting a significant link (12). That report included four case-control studies which examined both direct and SHS exposure; all found that active TB patients were more likely to be smokers (present or past) and/or have SHS exposure (25262728). In the one study that only examined SHS and active TB in adults, over 15,000 elderly, never-smoking, married Chinese women were followed prospectively; 117 cases of active TB were identified (29). The women with TB were more likely to have had SHS exposure than those without TB (29). While no studies have examined the frequency and amount of SHS exposure and its association with active TB, it appears that the intensity of CS exposure — as measured by number of cigarettes smoked per day, number of years smoked, or number of pack-years smoked — is significantly associated with active TB (2526273031323334353637). In 2 recent meta-analysis investigations, SHS was also significantly associated with active TB (1819) although the association with adults was not as great as that seen with children (18).
Limitations of epidemiologic studies linking SHS/direct CS exposure and TB
Epidemiologic studies can at best show an association between SHS/direct CS exposure and LTBI or active TB but cannot show any direct causal link. A potential confounder that may perhaps lead to increased TB transmission among those exposed to SHS and/or direct CS is congregation among smokers — leading to potential increase in TB transmission if there was a person with active TB in the group (12). Since cigarette smoking is generally associated with a lower socioeconomic status, with secondary link to malnutrition and crowded living conditions, these are additional confounders for acquiring TB (12). In studies of SHS exposure and TB contact investigation, it is not always clear whether SHS refers to the index TB case or the contacts. In either case, the active TB patient would be exposed to direct CS, SHS, or both and thus the increased association between SHS exposure and active TB/LTBI may be due to exacerbation of cough in the active TB subject from the SHS and/or direct CS exposure, resulting in greater transmission of Mycobacterium tuberculosis (MTB). Similarly, if indeed direct CS exposure is a risk factor for TB, those with SHS exposure may simply be at greater risk for TB because, by definition of having SHS exposure, they may be exposed more frequently to individuals (smokers) with active TB.
In summary, epidemiological studies show SHS exposure is associated with active TB and LTBI but such links are compromised by the great variability in the results (121819). Moreover, these studies do not show a cause-and-effect relationship and there is the issue of potential confounders such as malnutrition, crowded living conditions, and HIV co-infection. Even if there is a causal link between SHS/direct CS exposure and TB, epidemiologic studies cannot elucidate the mechanism(s). Thus, in the next 2 sections, we are going to: 1) tabulate the murine studies which investigated the effects of CS exposure (direct CS + SHS) on MTB infection and 2) summarize the main functions of key immune cells in control of MTB and the effects of SHS/direct CS on their functions.
MURINE STUDIES ON SHS/DIRECT CS AND TB
To the best of our knowledge, there have been three in vivo murine studies examining the effects of CS on control of MTB infection (383940). Each used CS exposure levels — based on total suspended particulate — that were comprised of both SHS and direct CS exposure. Each revealed that CS exposure impaired the ability of mice to control MTB infection (383940). We summarized the key findings of these mouse studies by immune cell types (Table 1).
Table 1
Effects of CS exposure on murine models of MTB infection

| Murine cell type | Findings in CS-exposed mice compared to air-exposed mice |
|---|---|
| Macrophages | Reduced number of IL-12 and TNFα-positive splenic and lung macrophages; increased number of IL-10-positive splenic macrophages (40). |
| Dendritic cells | Reduced number of IL-12 and TNFα-positive splenic and lung dendritic cells; increased number of IL-10-positive splenic dendritic cells (40). |
| T effector cells | Reduced number of TH1 cells with reduced IFNγ production per cell (383940); also reduced influx of IFNγ and TNFα-positive CD4+ and CD8+ T cells into the lungs and spleens (3840); increased influx of IL-4-positive TH2 cells in the lungs (39); CS also impaired (DNA) vaccinated mice to MTB-specific antigen by reducing the number of antigen-specific TH1 cells (38). |
![]()
THE EFFECTS OF SHS/DIRECT CS EXPOSURE ON IMMUNE CELLS RELEVANT FOR TB
CS significantly increases the number of innate and adaptive immune cells into the lungs (41) and yet epidemiologic evidence and experimental models indicate CS leads to a predisposition to TB (383940424344). This paradox supports the notion that the host-protective anti-MTB cell types are compromised and/or there is activation of immunosuppressive cells by SHS/direct CS. While CS extract impairs human macrophages in controlling MTB infection, the mechanisms remain unknown (4045). Hence, we highlight below some of the key immune cells relevant in host immune response to MTB and how SHS/direct CS may affect their functions.
Macrophage function against TB and potential effects of SHS/direct CS
M1 macrophage phenotype is characterized by production of pro-inflammatory cytokines (tumor necrosis factor [TNF] α, IL-12), chemokines IL-8 and monocyte chemoattractant protein-1, and nitric oxide as well as biasing T cells toward the TH1 phenotype and interferon (IFN) γ-producing CD8+ T cells (464748). M1 effector functions against MTB include phagosome-lysosome fusion, autophagy, apoptosis, efferocytosis, and granuloma formation (4950). Generally, M2 macrophages, the alternatively-activated or “deactivated” macrophage phenotype (in the context of host-defense against pathogens), are characterized by increased activity of arginase, matrix metalloproteinase (MMP), transforming growth factor (TGF) β, IL-10, and prostaglandin E2, and would not be expected to be host-protective during active MTB replication/infection (5152). Efferocytosis of MTB-infected neutrophils leads to macrophage activation through the actions of TNFα and heat shock proteins (53). Efferocytosis may assist in clearing MTB infection, enhance antigen presentation to T cells, limit potentially injurious inflammation, and increase phagosome-lysosome fusion through uptake of neutrophil azurophilic granules (5455). Table 2 lists the potential effects of SHS, direct CS, or nicotine on anti-TB macrophage function; whether murine cells (M), human cells (H), or both (M/H) were studied are noted for each of the studies cited.
Table 2
Potential effects of SHS/direct CS/direct or secondary nicotine on macrophage function against TB

| Macrophage phenotype and function | Effects of SHS/direct CS/direct or secondary nicotine |
|---|---|
| M1 phenotype | Skewed macrophages to the M2 phenotype (52)(H). Induced PD-L1/2, resulting in a deactivated phenotype (565758)(M/H,H,H). |
| Cell surface markers | Smoker AM had less expression of CD11a, CD71, and CD54; smoker AM have greater density of CD11c (but not percentage of AM positive for CD11c) (59)(H). |
| Phagocytosis | Effect of SHS/direct CS on phagocytosis is conflicting, from no effect to inhibition of phagocytosis (40456061)(M,H,H,H). |
| Efferocytosis | Inhibited efferocytosis (556263)(M/H,H,H) |
| Phagosome-lysosome fusion | Inhibited phagosome-lysosome fusion (646566)(R,H,M/H) |
| Autophagy | Inhibited autophagy by CS (5167)(M/H,H) and nicotine (68)(M/H). |
| Apoptosis | Inhibited apoptosis (6970)(H,M/H). |
![]()
Neutrophil function against TB and potential effects of SHS/direct CS
Lung neutrophils are one of the most commonly MTB-infected cells in TB patients and smokers have 3- to 4-fold more lung neutrophils than non-smokers (7172). Thus, whatever impact CS may have on neutrophils is likely relevant with MTB infection. Some studies show neutrophils restrict the growth or kill MTB whereas others show neutrophils contribute to excessive lung inflammation and may enhance mycobacterial dissemination (737475). Such discrepancies may be due to phenotypic differences in neutrophils as the CD16hiCD62Llo neutrophil subtype is considered immunosuppressive (76). Table 3 lists the potential effects of SHS or direct CS on anti-TB neutrophil function.
Table 3
Potential effects of SHS/direct CS on neutrophil function against TB

| Neutrophil phenotype and function | Effects of SHS/direct CS |
|---|---|
| Migration of neutrophils | Smokers have three- to four-fold more neutrophils in the lungs than non-smokers, likely due to CS induction of IL-8 and IL-17, the latter through CS differentiation of TH17 cells (7177)(H,M). |
| Neutrophil activation | PD-L1 is increased on neutrophils of active TB patients (78)(H) and CS, nicotine, and lipopolysaccharide (a common contaminant of cigarettes) are known to induce PD-L1/2 expression (5758)(M,H), theoretically deactivating neutrophils, leading to formation of non-protective, necrotic granulomas (75)(M/H). |
| Oxidative burst | CS inhibited endogenous oxidative burst (79)(H). |
| Induction of (N2) neutrophils | Lipopolysaccharide found in cigarettes induced the expression of immunosuppressive N2 (CD16hiCD62LloCD11bhiCD54hi) neutrophils, which inhibit T cell proliferation (76)(H). |
| NETs | CS inhibited endogenous oxidative burst — which is required for NETs formation; thus, CS could inhibit formation of NETs (7980)(H,M/H) |
![]()
T effector cell and B lymphocyte function against TB and potential effects of SHS/direct CS
Increasing evidence indicates that a TH1/TH2/TH17 balance is vital to establish control of active TB disease (81828384). MTB-specific TH1 cells (IL-12+, IFNγ+, TNFα+) stimulate inflammation, help skew macrophages to the M1 phenotype, and initiate antibacterial effector functions in infected macrophages (8586). Early expansion of TH2 cells (IL-4+, IL-13+) antagonizes protective cellular immunity, resulting in a partial or complete loss of TB control (468788). TH17 cells (IL-17+, IL-22+) recruit neutrophils through the chemoattractant actions of IL-17, resulting in improved early granuloma formation and increased cooperation between neutrophils and macrophages in killing MTB (89). However, sustained exposure of neutrophils to IL-17 can polarize their phenotype to one that is less effective in controlling MTB, causing more immunopathology (89). While increased programmed cell death protein 1 (PD-1) expression on T cells have been shown to impair human host immunity to MTB (90919293), several groups have shown that PD-1 knockout mice were paradoxically more susceptible to MTB, perhaps related to excessive inflammation and tissue injury (including neutrophil-mediated necrosis) in the complete absence of PD-1 (90949596).
CD8+ T cells are also required for optimal host defense against human MTB disease (4697). CD8+ T cells recognize protein and lipid antigens that are either class I MHC or CD1 restricted, respectively (9899). CD8+ T cells have the capability to recognize and kill MTB-infected macrophages (100101). Humans infected with MTB have expansion of cytotoxic CD8+ T cells capable of producing various cytokines (97). CD8+ T cells have also been identified in granulomas and pleural fluid of TB patients (102103104) and cloning these cells demonstrated TB antigen specificity (105). Additional reports demonstrated MTB-specific CD8+ T cells in BCG vaccinated individuals, and both active and latent TB patients (106107).
MAIT cells are a subset of CD8+ T cells that recognize bacteria-derived metabolites of riboflavin (vitamin B2) biosynthesis pathway that are presented on major histocompatibility complex-related protein-1 (MR1) located on the cell surface of antigen-presenting cells; interestingly, cell surface expression of MR1 is dependent on the presence of an exogenous, bacterial metabolite ligand (108). In contrast to conventional T cells, MAIT cells are functional prior to exposure to antigens although exposure to MR1-restricted microbial antigens induce MAIT cell expansion. The semi-invariant T cell receptors (TRAV1-2/TRAJ33) of MAIT cells that recognized MR1-restricted antigens are comprised of α and β subunits. When activated, MAIT cells are host-protective through the expression of IFNγ and TNFα, and may induce lysis of target cells through granulysin and perforin (109). In healthy humans, MAIT cells are already enriched in the respiratory tract but with active TB, the MAIT cell population is further increased in the lungs but decreased in the peripheral blood (108110111). But the MAIT cells in patients with active TB may be compromised as they exhibit increased expression of PD-1 and blockade of PD-1 signaling resulted in a significantly higher number of IFNγ-producing MAIT cells upon ex vivo stimulation with BCG (93).
There is increasingly convincing evidence that B cells — through production of antibodies but also by biasing specific CD4+ T cell phenotypic activation through cytokine production and antigen presentation to T cells — are an important component in an effective host immune response against MTB (112113114115116). While MTB is traditionally considered an intracellular pathogen, extracellular bacilli are a significant component of the total bacterial population. In this regard, there are at least 2 circumstances in which binding of specific antibodies to mycobacterial antigens optimizes antigen presentation; such optimization is perhaps required since many MTB cell wall components contain lipids and carbohydrates that are not typically recognized by MHC molecules. In 1 instance, mycobacterial antigens bind to cell surface antibodies on B cells, the immune complexes are internalized, and the antigens are processed and presented on class II MHC on B cells to T cells. Alternatively and/or concomitantly, free mycobacterial antigen-antibody complexes are able to bind to FcγR on the surfaces of professional antigen presenting cells (dendritic cells and macrophages) and be internalized. The antigens are then processed and presented to T cells. In turn, T cells are able to reciprocate and help B cells produce antibodies (117). In addition to enhancing antigen presentation, IgG can increase phagocytosis of MTB through increased binding of IgG-opsonized bacteria to FcγR or complement receptors present on macrophage cell surface, the latter occurring when C3 or C4 are bound directly to the mycobacteria or to IgG (114). Another potential antibody-mediated effector mechanism is opsonization of infected macrophages that express mycobacterial antigens on their cell surfaces with subsequent Fc binding to FcγRIII on cytolytic lymphocytes (114). In contrast to the active roles of B cells in anti-MTB immunity upon binding to stimulatory FcγR, binding of immune complexes to inhibitory Fc receptors such as FcγRIIB inhibits dendritic cell maturation, resulting in impaired T cell activation. Thus, antibodies produced by B cells may have both stimulatory and inhibitory effect on host immunity against MTB.
B and T cells are also able to reciprocally skew the differentiation of each other into specific phenotypes through the production of cytokines (112118). Remarkably, the specific subsets of B and T cells involved in this interaction are similar; i.e., B effector-1 cells (Be-1) produce IFNγ, IL-12, TNFα, IL-10, and IL-6 to skew naïve CD4+ T cells to the TH1 IFNγ+ phenotype and, in a reciprocal fashion, TH1 cells induce Be-1 differentiation. Similarly, Be-2 cells produce IL-2, lymphotoxin, IL-4, IL-13, IL-10, and IL-6 to induce TH2 IL-4+ differentiation and vice versa (112). Furthermore, B cell-derived TNFα serves to not only induce antibody production but also promote expansion of CD4+ T cell subsets, depending on the prevailing cytokine millieu.
Mice deficient in B cells or the common γ chain of FcγR are compromised in controlling an MTB infection, which may be due, in part, to increased IL-10 production (119120). Conversely, mice with genetic disruption of the inhibitory FcγRIIB have greater capacity to control MTB and have increased TH1 response, in part through increased IL-12 expression (120). In addition to the aforementioned host-protective mechanisms of B cells against MTB, they also contribute to the formation of acute and chronic granulomas in response to MTB (112121122123). In MTB infection of non-human primates, depletion of B cells with rituximab resulted in altered local T cell responses in granulomas (increased number of IL-2+, IL-10+, and IL-17+ T cells), decreased inflammation, and increased bacterial burden in individual granulomas although there was significant heterogeneity seen between the granulomas even in the same animal (124).
While B cells appear to play a lesser role than T cells in a respiratory mucosal vaccine against MTB (125), the humoral response also attenuates the potentially tissue damaging neutrophilic response, in part, by reducing the neutrophil chemokine IL-17 and the TH17 response (126). Indeed, B cell deficient mice have increased neutrophilia at the site of experimental immunization, with reduced dendritic cell migration to regional lymph nodes and decreased vaccine-induced TH1 response (126). As a promising therapeutic, monoclonal antibodies directed against various MTB cell wall components protected mice against MTB, as evinced by reduced bacterial load, containment from disseminated disease, and attenuated inflammation (127). Table 4 lists the potential effects of SHS or direct CS on anti-TB effector lymphocyte function.
Table 4
Potential effects of SHS/direct CS on effector lymphocyte function against TB

| Lymphocyte subtype | Effects of SHS/direct CS |
|---|---|
| TH1 cells | Reduced number of TH1 cells and decreased production of IFNγ per cell (128129)(H,H). Potential mechanisms include CS-derived reactive oxygen species-induced necrosis of T cells (130)(H), T cell exhaustion as CS increases PD-L1 on antigen presenting cells, leading to engagement of programmed death (PD-1) receptors on T cells (589192131)(H,H,H,M/H), downregulation of CD28, and upregulation of CTLA-4 (132)(M). In adolescents exposed to SHS, the percentages of blood memory CD4+ and CD3+ T cells (CD45RO+) were inversely related to the degree of SHS exposure although there was a direct relationship between the percentages of naïve CD3+ and CD4+ T cells (CD45RA+) and degree of SHS exposure (133)(H). In contrast, T cells from both CS-exposed mice and human smokers fluxed calcium, proliferated and produced IFNγ at levels comparable to T cells from control mice and non-smokers (134)(M/H). |
| TH2 cells | Increased influx of IL-4-positive TH2 cells in the lungs (39)(M). |
| TH17 cells | In mice, CS increased TH17 cells but only after relatively long-term exposure (135)(M). The percentage of TH17 cells is increased in the blood of healthy cigarette smokers (136)(H) and CS exposure of human lung explants increased IL-17 expression (137)(H). |
| CD8+ cells | Smokers have increased CD8+ cells in their airways than non-smokers (138)(H). CS increased the proliferation and inhibited the apoptosis of CD8+ T effector cells but increased both the proliferation and apoptosis of CD8+ Tregs (139)(H). CS exposure of mice increased CD8+ (and CD4+) T cells (140)(M). |
| MAIT cells | Human subjects exposed to CS had reduced circulating levels of CD26hiCD161hi MAIT cells (141)(H). |
| B cells | Chronic smoking is known to cause a polyclonal B cell lymphocytosis (142)(H). In adolescents exposed to SHS, there was no difference in the percentages of blood B lymphocytes and the degree of SHS exposure (133)(H). B cells from CS-exposed mice and human smokers fluxed calcium, proliferated and produced immunoglobulins at levels comparable to control mice and non-smokers (134)(M/H). In contrast, others have shown that CS-exposed or nicotine-exposed rodents have reduced number of B cells as well as reduced calcium flux and decreased proliferative capacity in response to antigenic stimulation although the amount of experimental CS exposure in these studies were greater than that typically seen in human smokers (143144)(M,M). |
![]()
Treg function against TB and potential effects of SHS/direct CS
Tregs secrete immunosuppressive cytokines such as IL-9, IL-10, IL-35, and TGFβ, increase consumption of IL-2, suppress Foxp3-negative TH1, TH2, and TH17 cells, and quell antigen-presenting cells (145146). Early in TB infection, excess Treg activity is associated with production anti-inflammatory of mediators (IL-10+, TGFβ+) that may promote MTB growth and persistence. In experimental animals, the influx of Tregs to the lungs with MTB infection worsens disease (147). Numerous studies have demonstrated that CD25+ and Foxp3+ Tregs are elevated in the circulation and at the site of infection in patients with active pulmonary TB (148149) and suppress the proliferation of and IFNγ production by CD4+CD25− T cells in an in vitro co-culture system (150). Conversely, in vitro depletion of CD25hi Tregs from peripheral blood mononuclear cells obtained from active TB patients resulted in further increase in IFNγ levels induced by specific antigens (151152). Other studies have confirmed these findings and determined that CD4+CD25+ Tregs in active TB co-expressed Foxp3, cytotoxic T-lymphocyte-associated antigen (CTLA)-4, glucocorticoid-induced tumor necrosis factor receptor (GITR), PD-1 and CD39 at the protein level (150153154) and also higher levels of CD45RO and human leukocyte antigen-antigen D related (HLA-DR) together with lower levels of CD127 (155) and CD45RA compared to CD4+CD25− T cells (154156157158). Patients with active TB have increased Tregs in their peripheral blood and bronchoalveolar lavage compared to subjects with latent TB infection or healthy controls (159). As would be predicted, co-culture of these isolated Tregs with T effector cells and MTB-infected monocyte-derived macrophages or alveolar macrophages significantly decreased mycobacterial killing (159). However, Treg function is also essential to control tissue damaging inflammation. Tregs express Foxp3+ CTLA-4 that have been linked to reduce TH17 cytokines such as IL-17 and IL-23 in subjects with LTBI (160161). Table 5 lists the potential effects of SHS, direct CS, or nicotine on Treg function.
Table 5
Potential effects of SHS/direct CS/direct or secondary nicotine on Treg function

| Tregs | Effects of SHS/direct CS/direct or secondary nicotine |
|---|---|
| Tregs | In man, CS exposure enhanced the function of Tregs in the lungs (162163164)(H,H,M). A possible mechanism by which CS enhances Treg function is the ability of CS to induce PD-L1/2 on antigen-presenting cells, resulting in increased engagement to PD-1 on Tregs, enhancing the immunosuppressive function of Tregs (5658165)(M/H,H,H). |
| In mice, Tregs express the α7 nicotinic acetylcholine receptor and upon binding to nicotine, there is an upregulation of both the transcription factor Foxp3 and the cell surface molecule CTLA-4, proteins that characterize Tregs and dampen Foxp3-negative T effector cell activity, respectively (164)(M). By producing TGFβ and IL-10, CS-induced Tregs can further suppress M1 macrophage activation against MTB as well as induce macrophages to differentiate to the M2/deactivated phenotype (51)(M/H). Nicotine induced murine Tregs to produce TGFβ and nicotine-exposed Tregs impair macrophage activity against MTB in murine macrophages (68)(M/H). |
![]()
CAN CS ENHANCE MTB VIRULENCE?
To the best of our knowledge, there has been no investigations to determine if CS influences the virulence of MTB. However, MTB is exposed to endogenous and exogenous reactive oxygen species — abundantly present in CS — and it is known that oxidative as well as reductive stressors may affect several physiologic functions of MTB, including intracellular signaling, synthesis of nucleic acids and proteins, and drug resistance (166). As in other host-pathogen interactions, MTB and host cells are likely altering its own redox balance to gain advantage over the other. McEachern and co-workers (167) showed in Staphylococcus aureus (S. aureus) that CS extract induced the expression of the multiple peptide resistance factor (mprF) protein, which adds lysine residues to acidic phospholipids (which have relative negative charge prior to lysine adducts) on the bacterial cell surface, converting them to basic phospholipids (relatively positive charge after addition of lysine residues), creating an unfavorable charge environment for the cationic antimicrobial peptide (AMP) LL-37 to bind bacteria, thereby generating a more virulent S. aureus (Fig. 1, left-side). Thus, it is highly relevant that MTB possess the lysX gene — a fusion gene comprised of both an mprF-like gene and the lysU gene — that encodes a protein with both lysyl transferase and lysyl-tRNA synthetase activities (168). A catalytic product of this 2-domain protein is lysinylated phosphatidylglycerol, a basic and positively-charged phospholipid, which imparts MTB resistance to cationic AMPs. Hence, it would be very interesting to determine whether CS could induce lysX gene expression, provoking greater MTB virulence through addition of lysine-mediated positive charge on MTB cell surface (Fig. 1, right-side).
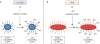 | Figure 1A hypothesized mechanism by which CS may affect virulence of MTB. (A) With S. aureus, CS induces the expression of mprF, which adds positively charged lysine residues to the bacterial membrane phospholipids. With greater positive charge on its cell membrane, S. aureus is better able to repel cationic AMP+. (B) Similar to the mprF molecule in S. aureus, MTB possess the lysX gene, which encodes a 2-domain protein with lysyl transferase and lysyl-tRNA synthetase activities that also adds positively charged lysine residues to phosphatidylglycerol, converting the acidic, negatively-charged phospholipid to a basic, positively-charged phospholipid. While it is not known if CS can induce lysX gene, this may be another potential mechanism by which CS increases susceptibility to TB by inducing a more virulent MTB strain.
|
SUMMARY
Both SHS and direct CS exposure are associated with increased risk for LTBI and active TB. This susceptibility may be due to increased exposure to the tubercle bacilli from the increased cough seen in smokers and those exposed to SHS, suppressed anti-TB immunity, enhanced activity of immunosuppressive N2 neutrophils or Tregs, or a combination of any of these factors. These generalized statements are likely an oversimplification of what is actually occurring since each cell type is not always salutorious or deleterious to the host; i.e., the temporal and coordinated influx (or absence) of each cell type may be necessary not only during the initial phase of the host-protective inflammatory response but also the necessary resolution of inflammation once the infection is under control. In addition, the potential effects of CS in inducing greater virulence in MTB remains to be determined but a plausibility given such findings in other bacteria. Finding stronger evidence of a causal link between SHS/direct CS exposure may provide greater impetus to implement public health policies to further reduce SHS/direct CS exposure in TB-endemic countries, potentially providing another important measure to help eradicate TB.
 XML Download
XML Download