

 PDF
PDF Citation
Citation Print
Print


Abbreviations
cLIA
competitive luminex immunoassay
E. coli
Escherichia coli
ELISA
enzyme-linked immunosorbent assay
GST
glutathione S-transferase
HPLC
high performance liquid chromatography
HPV
human papillomavirus
HRP
horseradish peroxidase
PAGE
polyacrylamide gel electrophoresis
PBNA
pseudo virion-based neutralization assay
PBST
PBS with 0.1% tween
RT
room temperature
VLP
virus-like particle
INTRODUCTION
Human papillomavirus (HPV) is a member of the papoviridae virus and consists of a small double-stranded DNA virus containing a circular genome of approximately 8,000 base pairs without an envelope (1). HPV infection is the primary cause of cervical cancer, which can be sexually transmitted (2). The sequence variation on L1 protein, the major element of the viral capsid, encoding gene defines the types of HPV. Over 40 types of different HPVs are responsible for the anogenital, cervical, and oropharyngeal epithelia infections (3). Low-risk subtypes such as HPV type 6, 11, 42, 43, and 44, lead to warts and condyloma acuminata (genital warts) while the high-risk subtypes such as HPV type 16, 18, 31, 33, 34, 35, 39, 45, 51, 52, 56, 58, 59, 66, 68, and 70, have been shown to be the cause of cervical cancer (4). HPV type 16 is the cause of about 54% of invasive cervical cancers and HPV type 18 is known for the 17% of worldwide HPV infection (5). Furthermore, HPV type 6 or 11 infections are associated with approximately 90% of all genital warts (6).
HPV expresses 6 early open reading frame (ORF) proteins, E1, E2, E4, E5, E6, and E7 with regulatory functions beside 2 late ORF proteins, major capsid protein L1 and minor capsid protein L2 (7). The self-assemble of major capsid protein L1 into virus-like particles (VLPs) is highly immunogenic (8). L2 protein co-assembles with L1 to form VLPs at the same 1:30 molar ratio as that found in authentic virions (9). Electron photo-micrograph analysis shows that L1 and L1/L2 VLPs cannot be morphologically distinguished from infectious virions, except the lack of electron-dense core that is characteristic of encapsidated viral genome (10). It was confirmed that after parenteral injection L1 VLPs can induce the high titers of serum antibodies that neutralize bovine papillomavirus type 1 in vitro (11) and similar data were obtained with oncogenic HPVs (121314).
A quadrivalent HPV recombinant vaccine has been developed against HPV types 6, 11, 16, and 18. This vaccine contains the VLPs derived from L1 capsid proteins of HPV type 6, 11, 16, and 18 expressed in Saccharomyces cerevisiae. Heterogenic eukaryotic expression systems have recently been employed to generate VLPs (15). The recombinant VLPs proteins have been produced in animal cells (16), insect cells (17), yeast (18), as well as bacterial cells (19) and used the leading candidate vaccines to prevent cervical cancer (20).
HPV competitive radioimmunoassay (cRIA) (21), HPV competitive luminex immunoassay (cLIA) (22), and enzyme-linked immunosorbent assay (ELISA) (23) using VLPs as antigens have been used for quantification of the quadrivalent HPV vaccine. The cLIA assay engages fixing 4 distinct VLPs of HPV type 6b, 11, 16, and 18 to Luminex microspheres with different fluorescent properties (24). The ability of sera samples derived from vaccines to prevent VLP binding is assessed by a type-specific neutralizing monoclonal antibody, which has a phycoerythrin fluorescent tag. For cRIAs HPV type 6b, 11, 16, and 18 L1 VLP antigens coated polystyrene beads are incubated with the compatible HPV type 6b, 11, 16, or 18 specific mouse antibodies (mAbs) and serum sample. 125I-tagged goat anti-mouse antibodies were used for detection of mAbs bound on VLPs (2526). VLP ELISA was developed firstly for HPV type 16 (27), and later this technique has been developed for other HPV types (28). In addition, HPV L1 proteins (type 6b, 16, and 18) fused to glutathione S-transferase (GST) was used for GST based ELISA which uses glutathione cross-linked to casein as capture protein to purify and immobilize bacterially expressed GST-L1 (27) fusion proteins as antigens. Pseudovirion-based (829) neutralization assay (PBNA) was used to measure a range of functional neutralizing antibodies although ELISA was used to measure both neutralizing and non-neutralizing antibodies.
The present study demonstrated the production of viral capsid protein L1 of HPV type 11, 16, and 18 expressed in Escherichia coli (E. coli) as N- and C-terminal proteins. Furthermore, titration assay and western blot techniques with vaccinated and non-vaccinated human sera were used to observe whether there is any reactivity by HPV vaccine.
MATERIALS AND METHODS
Molecular cloning of the HPV L1 protein encoding genes
HPV type 11, 16, and 18 DNA derived from ATCC Company (Manassas, VA, USA) were amplified by PCR, 2 different reaction mixes prepared for each virus type to obtain N- and C-terminal L1 protein fragments. PCR amplified DNA fragments were purified using RBC Bioscience genomic DNA extraction kit (Taipei City, Taiwan). Purified fragments were ligated with pProEx/HTa vector except the C-terminal HPV type 16 coding gene which was ligated with pProEx/HTb vector as described (30313233). After the transformation of the cells into E. coli DH5α strain, the cells were spread on ampicilin (100 µg/ml) containing LB Agar plate and incubated overnight. The colonies harboring ligated constructs were screened by PCR, the plasmid DNAs were purified using plasmid miniprep kit from Cosmo Genetech (Seoul, Korea). The successfully ligated plasmid constructs were sequenced by Cosmo Genetech (Seoul, Korea).
Recombinant N- and C-terminal L1 protein expression and purification
N- and C-terminal L1 protein expressing gene containing DH5α clones were incubated in 10 ml of LB media containing ampicillin (100 µg/ml) at 37°C overnight. The overnight culture was inoculated into 500 ml of LB medium and grown till the cells reach mid-exponential stage (OD value of 0.6–0.7 at A600). The protein expression was induced by adding 0.1 mM of isopropyl β-D-1-thiogalactopyranoside (IPTG) incubating the cells at 37°C for 4 hours. The induced cells were pelleted down by centrifugation (6,000 rpm, 4°C, for 20 min). After resuspension of the cells in appropriate buffers which were shown in Table 1, suspension was pulse-sonicated at 10 s intervals for 1 min. In the case of urea containing buffers suspensions were incubated at 37°C overnight and of guanidine buffer at 4°C for 4 h.
Table 1
The list of lysis buffers was used for protein extraction

![]()
Following the incubation step sonication was performed one more time. The protein solution was centrifuged at 10,000 rpm for 10 min (for urea buffer at 25°C and for guanidine buffer at 4°C). After the centrifugation the proteins in supernatant was purified with a TALON affinity column (Invitrogen, Carlsbad, CA, USA) using his-6x-tag at the N-terminal of recombinant proteins. The TALON affinity-purified proteins were subjected to high performance liquid chromatography (HPLC) from Agilent Technologies (Santa Clara, CA, USA) as described (3435363738).
Serum titration assay
The 96-well immunoplate (SPL Life Sciences, Pocheon, Korea) was coated with successfully purified N- and C-terminal L1 protein of HPV type 11, 16, and 18 (1 µg/ml) and incubated at 4°C overnight. Next day 200 µl of blocking solution (2% bovine serum albumin [BSA] and 0.02% NaN3 in PBS) was added and 96-well plate was incubated at room temperature (RT) for 1 hour. After blocking, the plate was washed with 200 µl of PBS with 0.1% tween (PBST). This study was approved by multiple institutional review board (Seoul, Korea). Vaccinated and non-vaccinated human sera were obtained from healthy individuals that were added on protein coated wells with 1:300, 1:600, and 1:1,200 dilution ratios. Following 2 hours incubation at RT the plate was washed with PBST for 3 times. Goat anti-human IgG conjugated with horseradish peroxidase (HRP) was diluted in PBS and added on wells (100 µl). After incubation at RT for 40 min, the plate was washed with PBST for 3 times then 3,3′,5,5′-tetramethylbenzidine (TMB) solution (100 µl) was used for developing (Thermo Scientific, Rockford, IL, USA).
Western blot analysis
The SDS-polyacrylamide gel electrophoresis (PAGE) gels were electro-transferred onto nitrocellulose membranes (GE Healthcare, Pittsburgh, PA, USA) for 50 min at 0.3 mA using Bio-Rad transblot apparatus. The membranes were blocked in 5% skim milk in PBST (3.2 mM Na2HPO4, 0.5 mM KH2PO4, 1.3 mM KCl, 135 mM NaCl, 0.05% Tween-20, pH 7.4) for 20 min at RT. Then the membranes were incubated with human sera (1 to 1,000 dilution in blocking solution) overnight at 4°C. After incubation the membranes were washed with PBST 3 times in a row, each time for 5 min. Goat anti-human IgG HRP conjugate was diluted in blocking solution and membranes were incubated in this mix for 40 min at RT.
RESULTS
Molecular cloning and expression of HPV L1 capsid protein encoding genes
The N- and C-terminal L1 protein of HPV type 11, 16, and 18 were amplified by PCR from a sample obtained from ATCC Company. The primer sets introduced EcoRI (by forward primer) and XhoI (by reverse primer) restriction sites. After the restriction and purification of PCR amplicon and the vectors pProEx/HTa and pProEx/HTb, N-terminal of all 3 types of HPV, C-terminal L1 protein of HPV type 11 and 18 were cloned into pProEx/HTa and in the case of C-terminal L1 of HPV type 16 into pProEx/HTb. The cloned plasmid DNAs were confirmed both by PCR and DNA sequencing (data not shown).
L1 protein of the HPV species HPV type 11, 16, and 18 were expressed in 2 parts as N- and C-terminal. According to the small-scale expression results N- and C-terminal L1 protein is expressed with higher levels in E. coli strain Codon plus or Rosetta comparing to DH5α (Fig. 1), yet expression of HPV type 11 N-terminal L1 protein is expressed highly in DH5α as well. In all cases protein is found in the insoluble fraction.
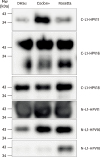 | Figure 1Small scale expression of HPV L1 proteins. C- and N-terminals of HPV-L1 protein were expressed in different E. coli strains: DH5α, Codon+, and Rosetta. The strain with highest protein yield was chosen for further steps. The data represents one of 3 independent experiments.
|
N- and C-terminal of L1 protein coding sequences were cloned into pProEx/HTa vector and pProEx/HTb in the case of C-L1-HPV type 16. During the large-scale protein production expression was induced by IPTG. After the TALON purification SDS-PAGE was performed and N-terminal L1 protein of HPV type 11 and C-terminal L1 protein of HPV type 16 gels' Coomassie blue staining results were shown in Fig. 2A and C, yet C-terminal L1 of HPV type 11 and 18 had relatively lower yield, therefore western blotting images were shown (Fig. 2B and D). Moreover, a second band was observed for TALON column purified C-L1-HPV type 16 and 18 on the SDS-PAGE gel. As next step HPLC was performed and the purified proteins were screened by silver staining (Fig. 3). After HPLC small C-L1-HPV type 16 protein fragments appeared on the silver stained gel. Nevertheless, the production of N-terminal L1 protein of HPV type 16 and 18 is failed (data is not shown).
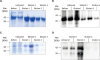 | Figure 2Expression of HPV L1 proteins in large scale. Large scale expression of (A) N-terminal L1 HPV type 11, (B) C-terminal L1 HPV type 11, (C) C-terminal L1 HPV type 16, and (D) C-terminal L1 HPV type 18. Coomassie blue staining and western blotting results after TALON purification were shown. The data represents 1 of 4 independent experiments.
|
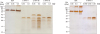 | Figure 3Further purification and quantitation of recombinant L1 proteins. The first step TALON chromatography purification of HPV-L1 proteins were further purified with HPLC. The purities and concentrations of HPV L1 proteins were examined by silver staining compared to BSA. Each recombinant protein was indicated on the top. The data represents 1 of 4 independent experiments.
BSA, bovine serum albumin.
|
Titration assay and western blotting to see the antibody response after vaccination
Serum titration assay was performed to see whether there was antibody production against HPV after vaccination, by using non-vaccinated and vaccinated human sera. However, no difference detected between the vaccinated and non-vaccinated samples, except HPV type 11. Both N- and C-terminal HPV type 11 L1 protein showed higher signals with vaccinated human sera comparing to non-vaccinated ones (Fig. 4A). In addition, C-terminal HPV type 16 and 18 showed much higher signals as a result of assays both for vaccinated and non-vaccinated human sera comparing to other antigen types (Fig. 4B).
 | Figure 4Serum titration of the quadrivalent HPV vaccine. (A) The data was showing the titration assay results using N- or C-terminal L1 protein of HPV type 11 as antigen and (B) showing the results using C-terminal L1 of HPV type 16 or type 18 as antigen. Representative data from 1 of 3 independent experiments are shown.
SEM, standard error of mean.
Mean SEM (n=7); *p<0.05 from 3 replicates.
|
To investigate if the HPV vaccine Gardasil (Merck, NJ, USA) induces antibody production after appropriate vaccination (3 times), human sera derived from vaccinated and non-vaccinated individuals, and healthy individuals were used for the western blot analysis. Interestingly it has been seen that 2 of the vaccinated human sera out of 4 possessed antibody against N-terminal HPV type 11 (Fig. 5A). Once the 2 sera samples from non-vaccinated humans were compared with sera of same individuals but after vaccination an antibody response against N-terminal L1 HPV type 11 was recognized (Fig. 5B and C).
 | Figure 5Western blot analysis using N-terminal L1 HPV type 11. (A) Western blot was performed by using N-terminal L1 proteins of HPV type 11 with sera of quadrivalent HPV vaccinated individuals as primary sera; (B) non-vaccinated individuals' sera did not contain antibody against N-terminal L1 protein of HPV type 11; and (C) while N-terminal HPV type 11 was detected by the same individuals' sera after 3 times injection with the HPV vaccine.
|
Moreover C-terminal L1 protein of HPV type was captured in western blot analysis in both cases with vaccinated human sera and non-vaccinated human sera (Fig. 6), yet on the further steps it was shown that binding on the C-terminal L1 protein of HPV type 16 and 18 were non-specific by using the control samples of human sera derived from healthy individuals in both males and females (Fig. 7).
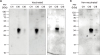 | Figure 6Western blot analysis to detect C-terminal L1 HPV type 11, 16, and 18 using sera of vaccinated and non-vaccinated individuals. The purified C-terminal L1 proteins of HPV type 11, 16, and 18 were used to perform western blot with human sera from vaccinated (A) and non-vaccinated (B) individuals. The data represents one of 3 independent experiments.
|
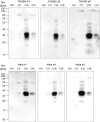 | Figure 7Control experiment using male and female sera samples form healthy individuals. Western blot is performed with serum derived from healthy individuals' sera as control. Binding with C-terminal L1 HPV type 16 and 18 were detected from both male and female sera. The data represents one of 3 independent experiments.
|
However, one of the samples, which were derived from HPV vaccine injected healthy female, captured N-terminal L1 of HPV type 11 as well. To confirm that C-L1-HPV type 16 and 18 binding was non-specific, western blot was repeated using only secondary antibody, but not human sera and it was reported that the L1 proteins had an interaction with anti-human IgG Fc HRP (Fig. 8).
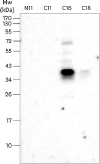 | Figure 8Control experiment showing non-specific binding of C-terminal L1 HPV type 16 with α-human IgG HRP. Western blot is performed using N-terminal L1 HPV type 11, and C-terminal L1 protein of HPV type 11, 16, and 18. Instead of using human sera as primary antibody, anti-human IgG HRP was used directly to detect the non-specific binding. The data represents one of 3 independent experiments.
|
DISCUSSION
We have expressed in E. coli and purified N-terminus L1 protein of HPV type 11 and C-terminus L1 of HPV type 11, 16, and 18. All proteins were insoluble, but because of their different characteristics we used different lysis buffer listed in Table 1 for each of the proteins. As reported by the previous studies the HPV L1 proteins were expressed in bacteria, yet the product was incorrectly folded protein with less immunogenicity than properly assembled particles (29). Since we divided L1 protein as N- and C-terminus, we aimed to overcome this problem. Yet we failed to express the N-terminal L1 protein of HPV type 16 and type 18. This result could be explained by the inhibitor function of N-terminal of HPV16 L1 protein, which has been shown by Ma et al. (39).
By using non-vaccinated, vaccinated and healthy human sera titration assay is performed to examine the antibody forming of vaccine, yet no significant signals are detected, except N-terminal L1 of HPV type 11 as well as C-terminal. To see the reason behind the high signals from the samples using C-terminal L1 HPV type 16 and 18 as antigens, we did further testing with healthy human sera and additionally without sera but the secondary antibody, as a result the binding of these antigens were non-specific.
To support the titration assay carried out with vaccinated and non-vaccinated human sera, we performed western blot. According to the western blot results the vaccinees' sera comprise antibody against N-terminus L1 protein of HPV type 11. Moreover, after the vaccination of initially not vaccinated individuals, similarly to our previous data, we observed a response to N-terminal L1 of HPV type 11. On the other hand, the C-terminal recombinant proteins of HPV type 16 and 18 bind with anti-human IgG which we used as secondary antibody conjugated with HRP both for ELISA and western blot analysis.
There are several assays to screen antibody responses to HPV, including PBNA, ELISA, cLIA, and the in situ-purified GST L1 fusion protein based ELISA (21222340). However, there is no standardized serological assay available for the evaluation of HPV-specific antibody responses. Although the PBNA is called as golden standard in HPV serology, this assay is highly labor intensive and therefore difficult to apply in large epidemiological studies. The cLIA measures antibodies directed against only one neutralizing epitope for a distinct HPV type, which can result in an underestimation of total neutralizing activity of HPV-specific antibodies. GST-L1 ELISA and VLP-based ELISA measure both antibodies against conformational and linear epitopes, thereby detecting all HPV-specific antibodies. It has been shown that the VLP-based ELISA correlates well with the PBNA (4142). Each assay quantifies an overlapping but distinct subset of only HPV type 16/18 VLP antibodies.
Another complication of the quadrivalent HPV vaccine is it only consists of HPV major capsid protein L1 but not the minor L2. As reported by Hernandez et al. (43) HPV L1 VLP and L1–L2 VLP serologic assays to measure virion antibodies to HPV type 16 and 18 simultaneously in a multiplex format were developed. Similar results were observed for both seroassays done with HPV type 16 yet concordance for HPV type 18 was much lower. It is suggested that HPV type 18 L1 VLP seroassay detects non-neutralizing antibodies and cross-reactive antibodies while L1–L2 VLP assay detects type-specific neutralizing antibodies. An alternative statement is that L1–L2 capsid is more stable than L1 capsid.
The antibody forming of the quadrivalent HPV vaccine was not sufficient to detect the antigens although the present study was performed with ELISA and western blot with few limited HPV antigens. The experiments were performed using HPV-L1 antigens failed to show the antibody response after vaccination with all the antigens produced. It is necessary to express robust recombinant L1 major capsid protein of all HPV types (6b, 11, 16, and 18) to investigate antigen forming by quadrivalent HPV vaccine. Further study using larger number of samples with biochemical and molecular level of analysis may help to understand specific antibody forming after quadrivalent HPV vaccine.
 XML Download
XML Download