

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
2-D
2-dimensional
2-DE
2-dimensional electrophoresis
Akt
protein kinase B
BCR
B cell receptor
GSK3β
glycogen synthase kinase 3β
hnRNP
heterogeneous nuclear ribonucleoprotein
LPS
lipopolysaccharide
MAP
mitogen-activated protein
MHC
major histocompatibility complex
p
phosphorylated
PDK1
3-phosphoinositide-dependent protein kinase 1
PI3K
phosphoinositide 3-kinase
RQ
relative quantitation
INTRODUCTION
The development of B cells from progenitor cells into terminally differentiated plasma cells is regulated by a carefully orchestrated network of signal transduction and the ordered expression of a large number of genes. Among the signaling network components, mitogen-activated protein (MAP) kinase cascades play an important role in the regulation of mammalian cell proliferation in a manner inextricable from other signal transduction systems by sharing substrates and cross-cascade interactions (1). Specifically, p38 and JNK MAP kinases, which can be activated by cellular stress, are closely related to B cell survival and activation via B cell receptor (BCR), CD40, and B-cell activating factor activation (2). In addition, p38 directly phosphorylates and eventually establishes myocyte-specific enhancer factor 2C as a direct transcriptional effector of BCR signaling in B cell proliferation (3). The protein kinase B (Akt) signaling pathway is another important signaling pathway of B cell activation that mediates various biological responses, including inhibition of apoptosis and stimulation of cell proliferation (4). Following activation, Akt phosphorylates a large number of downstream effectors, including mouse double minute 2 homolog, glycogen synthase kinase 3β (GSK3β), forkhead box O3, Bcl-2-associated death promoter, caspase-9, p27, and tuberous sclerosis complex 2, resulting in B cell growth, survival, and proliferation (5).
The major histocompatibility complex (MHC) class II molecule, a heterodimeric cell surface glycoprotein, is expressed on various Ag-presenting cells (APCs), including dendritic cells (DCs), phagocytes, and B cells (6). Although the MHC class II molecule is primarily known for its Ag-presenting function, its role in the homeostatic regulation of lymphocytes is well-documented. Although MHC class II molecules lack any known signaling motifs in their short cytoplasmic tail, it has been proposed that associated signaling molecules at the membrane or inside the cell mediate signaling via MHC class II molecules (7). For example, intracellular MHC class II molecules act as adaptors to promote full activation of the Toll-like receptor (TLR)-triggered innate immune response in macrophages by activating Bruton's tyrosine kinase (Btk) (8). In several mouse models, altered intracellular homeostasis of MHC class II molecules, including transgenic overexpression of MHC class II molecules and knockout of CD74 or MHC class II α-chain, is responsible for a concurrent loss of B cells (9). In addition, B cells from signal peptide peptidase-like protein 2-MHC class II double-deficient mice are defective in B cell maturation (10). We previously reported a function of MHC class II molecule signaling in B cell development whereby ligation of MHC class II molecules to a cognate anti-MHC class II Ab inhibited lipopolysaccharide (LPS)-stimulated B cell proliferation and differentiation; however, it is still unclear which signaling molecules are involved in the MHC class II-mediated inhibitory effect on B cell proliferation and differentiation (11).
To understand the characteristics of MHC class II molecule-mediated signaling, we identified and co-immunoprecipitated various candidate proteins associated with MHC class II molecules and showed differential protein expression patterns in LPS-stimulated 38B9 B cells with versus those without anti-MHC class II Ab treatment. Among them, we selected heterogeneous nuclear ribonucleoprotein (hnRNP) A2B1 and investigated its function in relation to cellular responses. In this study, we investigated the role of hnRNP A2B1 in cellular signal transduction involved in LPS-induced B cell survival, proliferation, and differentiation in 38B9 B cells.
MATERIALS AND METHODS
Chemicals and laboratory wares
Unless otherwise specified, the chemicals and laboratory wares used in this study were obtained from Sigma Chemical Co. (St. Louis, MO, USA) and SPL Life Sciences (Pocheon, Korea), respectively. Oligonucleotide primers and nucleotide sequencing were both provided by Bioneer Inc. (Daejeon, Korea).
Cell culture
The pro-B cell line 38B9, established from BALB/c mice (d-haplotype) and with a μ heavy chain-negative and surrogate light chain-positive phenotype, was cultured in RPMI-1640 medium (Welgene, Gyeongsan, Korea) supplemented with 10% fetal bovine serum (GE Healthcare Life Sciences, Pittsburgh, PA, USA) and 50 μM 2-mercaptoethanol at 37°C in a humidified CO2 incubator.
Immunoprecipitation
MHC class II-associated protein complexes were prepared using an immunoprecipitation procedure described previously (12). In brief, cell lysates were precleared by incubating with a slurry of protein A-Sepharose 4B (GE Healthcare Life Sciences) with rotation for 30 min. The sample was then centrifuged at 5,000 g for 3 min, and the pellets were discarded. The collected supernatant was mixed and incubated with MK-D6 anti-I-Ad Ab with continuous rotation for 4 h at 4°C. A slurry of protein A-Sepharose 4B was added, and the mixture was incubated overnight at 4°C. The beads were then washed with lysis buffer before resuspension in sample reducing buffer.
Two-dimensional electrophoresis (2-DE)
The 38B9 cells that had been treated with LPS or LPS together with MK-D6 were lysed in lysis buffer. Each immunopellet was collected by immunoprecipitation and freeze-dried. Freeze-dried proteins were precipitated using the 2-dimensional (2-D) Clean-up Kit (GE Healthcare Life Sciences) before resuspension in rehydration buffer. The protein concentration was determined using the RC/DC Protein Assay Kit (Bio-Rad, Hercules, CA, USA). Samples were loaded onto the strip holder, covered with an Immobiline Drystrip (GE Healthcare Life Sciences) and rehydrated passively at 20°C for 12 h. Isoelectric focusing was performed for a total of 20,000 V/h using a linear ramp protocol at room temperature. The strips were then equilibrated for 15 min each with buffer I and buffer II. The strips were loaded on top of the gels, and a 2-D run was performed at 70 V for 2 h. The gels were stained with Coomassie brilliant blue, and stained gels were analyzed using PDQuest™ 2-D analysis software (Bio-Rad).
Mass spectrometry
Mass spectrometry analysis was performed as described previously. In brief, protein spots excised from the 2-D gel were dehydrated and dried in a concentrator. The gel pieces were then reduced with dithiothreitol and alkylated with iodoacetamide. After washing, gel pieces were swollen in digestion buffer containing sequencing-grade trypsin (Promega Co., Madison, WI, USA). Solutions containing peptides released into the buffer were collected as follows. Gel pieces were extracted twice with 0.1% trifluoroacetic acid in water for 20 min, and the soluble fractions were pooled together and dried. The final pellet contained most of the tryptic peptides from the digest and was analyzed by tandem mass spectrometry (MS-MS) using a Quadrupole Time-of-Flight (Q-TOF) mass spectrometer (QSTAR® XL Mass spectrometer, Applied Biosystems/MDS Sciex, Foster City, CA, USA). Protein identification using the generated data was performed using MDS Sciex, and molecular masses and isoelectric points were calculated using the web-based ExPaSy computer molecular weight/isoelectric point tool.
siRNA transfection and sample preparation
Synthesized siRNA targeting hnRNP A2B1 (5′-GGA UUU GGC UUU GUA ACU U-3′) and a negative control siRNA (5′-UUC UCC GAA CGU GUC ACG UTT-3′) were obtained from Genepharma Co., Ltd. (Shanghai, China). To measure the level of cell signaling, 38B9 cells (105) were transfected with an siRNA mixture consisting of 400 nM hnRNP A2B1 siRNA or negative control siRNA using Lipofectamine 3000 (Thermo Fisher Scientific, Waltham, MA, USA). Transfected cells were incubated in a CO2 incubator at 37°C. The cells were used for subsequent analysis at 24 h after transfection. After transfection, harvested cells were treated with 25 μg/ml LPS at 37°C for 30 min. The cells were then used for RNA or protein extraction.
To measure cytokines and cell maturation, 38B9 cells were transfected with an siRNA mixture consisting of 400 nM hnRNP A2B1 siRNA or control siRNA using Lipofectamine 3000 (Thermo Fisher Scientific). Transfected cells were incubated in a CO2 incubator at 37°C for 12 h and then treated with 25 μg/ml LPS for 12 h. To measure cytokine levels in the culture supernatants, the culture supernatants were collected 12 h following LPS treatment. The levels of various cytokines in the collected supernatants were determined using the Th1/Th2/Th17 BD Cytometric Bead Array Kit (BD Pharmingen, Franklin Lakes, NJ, USA). Harvested cells were fixed in 2% paraformaldehyde and subjected to cytometric analysis using anti-mouse IgM Ab followed by incubation with a FITC-conjugated secondary Ab or used for quantitative real-time PCR.
RNA isolation and quantitative real-time PCR analysis
The expression levels of various genes were determined using RT-PCR and quantitative real-time PCR. Total cellular RNA was isolated using the Easy-BLUE™ Total RNA Extraction Kit (Intron Biotechnology, Sungnam, Korea), and prepared RNA was converted into cDNA using a reverse transcription reaction system (Promega Co.) with a random primer or Jκ2 primer sets. PCR amplification was performed using a reaction mixture containing 10 pmol each of forward and reverse primers and cDNA (13). The PCR products were analyzed by electrophoresis on a 1.5% agarose gel. Quantitative real-time PCR was carried out in a 96-well plate using gene-specific primer sets and the Quantitect SYBR Green PCR Kit (Qiagen, Hilden, Germany) on the ABI 7500 system (Applied Biosystems/MDS Sciex). The expression level of the germline κ gene was normalized to that of actin via a relative quantitation (RQ) method using 7500 FAST software version 2.0.6 (Applied Biosystems/MDS Sciex).
The gene-specific forward and reverse primer sets used to amplify each gene were as follows: 1) hnRNP A2B1-F: 5′-CCG ATA GGC AGT CTG GAA AG-3′, hnRNP A2B1-R: 5′-CAT AAT TTC CTC CTC CAT AG-3′; 2) GAPDH-F: 5′-CCG ATG CCC CCA TGT TTG TG-3′, GAPDH-R: 5′-GGC CAT GCC AGT GAG CTT CC-3′; 3) actin-F: 5′-CGT ACC ACA GGC ATT GTG-3′, actin-R: 5′-CTC GTT GCC AAT AGT GAT GA-3′; and 4) germline Igκ-F: 5′-CCG GAT CCA CGC ATG CTT GGA GAG GGG GTT-3′, germline Igκ-R (Jκ2): 5′-CCA AGC TTT CCA GCT TGG TCC CCC CTC CGA A-3′.
Western blot analysis
For western blot analyses, cells were washed with cold phosphate-buffered saline and then lysed in mammalian protein extraction reagent (Thermo Fisher Scientific) or NE-PER nuclear and cytoplasmic extraction reagents (Thermo Fisher Scientific) supplemented with a protease inhibitor cocktail (Thermo Fisher Scientific) and a phosphatase inhibitor (Roche, Mannheim, Germany). Protein concentration was determined using a commercial BCA system (Thermo Fisher Scientific) with bovine serum albumin (BSA) as a standard. The lysates were resolved by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Millipore, Billerica, MA, USA). The membranes were blocked with 3% BSA/Tris-buffered saline containing Tween 20 and incubated first with specific Abs and then with a horseradish peroxidase-conjugated secondary Ab. Finally, the blots were developed using the enhanced chemiluminescence detection system (GE Healthcare Life Sciences).
Abs against hnRNP A2B1, ERK, p38, JNK, actin, GAPDH, p50, p52, c-Rel, and RelB were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). Abs against Akt, phosphorylated (p)-Akt (Ser), p-Akt (Thr), 3-phosphoinositide-dependent protein kinase 1 (PDK1), p-PDK1 (Ser), p-ERK, p-p38, p-JNK/stress-activated protein kinase (SAPK), and Mcl-1 were purchased from Cell Signaling Technology (Danvers, MA, USA).
Statistical analysis
Statistical analysis was performed using Prism 5 (GraphPad Software, La Jolla, CA, USA). Data are presented as means±standard deviation of repeated experiments. The unpaired Student's t-test was used to compare 2 groups. Differences in mean values were considered significant at p<0.05.
RESULTS
The recruitment of hnRNP A2B1 is inhibited by anti-MHC class II Ab treatment in LPS-treated 38B9 cells
We previously reported that LPS-mediated activation of 38B9 B cells was suppressed by crosslinking of MHC class II molecules via anti-MHC class II Ab treatment (14). To monitor the change in MHC class II molecule-associated proteins during this inhibition, MHC class II molecule-associated proteins were separated by 2-DE and analyzed using Q-TOF mass spectrometric analysis. We identified 10 candidate proteins showing enhanced expression after LPS stimulation and decreased expression after anti-MHC class II Ab treatment. Among them, we selected hnRNP A2B1, which regulates the transcription of genes involved in cell cycle progression and cell proliferation (15). Initially, we investigated whether hnRNP A2B1 is involved in MHC class II molecule-associated cell signaling (Fig. 1). The intensity of the spot corresponding to hnRNP A2B1 on the 2-DE gel suggested that LPS treatment of 38B9 cells increased the recruitment of hnRNP A2B1 to MHC class II molecules; this enhancement was inhibited by co-treatment with cognate anti-MHC class II Ab, which was confirmed at the hnRNP A2B1 transcript level (Fig. 1A). We detected similar enhanced expression of hnRNP A2B1 after LPS treatment and anti-MHC class II Ab-mediated inhibition of the enhanced hnRNP A2B1 expression in 38B9 cell lysates (Fig. 1B). Finally, we confirmed the direct interaction between hnRNP A2B1 and MHC class II molecules by immunoprecipitating MHC class II molecules. The level of MHC class II molecule-associated hnRNP A2B1 was measured in the immunoprecipitates and found to be increased by LPS treatment; the enhanced level of hnRNP A2B1 was reduced by anti-MHC class Ab treatment (Fig. 1C). Collectively, we confirmed that hnRNP A2B1 is associated with MHC class II molecules and involved in activation signaling in 38B9 B cells.
 | Figure 1Identification of hnRNP A2B1 as an MHC class II-associated protein in 38B9 cells. (A) Partial 2-DE image of hnRNP A2B1 (upper panel) and the RT-PCR product of hnRNP A2B1 mRNA (lower panel) prepared from 38B9 cells in the presence or absence of LPS and anti-MHC class II Ab. MHC class II-associated proteins were prepared by immunoprecipitation from whole-cell lysates using an anti-MHC class II Ab and separated by 2-DE as described in the Materials and Methods section. GAPDH was used as an internal loading control for RT-PCR. Molecular mass and isoelectric point of the candidate protein included in the spot were calculated as 37.4 kDa and 8.97, respectively, using AxPASy inline tool and candidate protein was expected to be hnRNP A2B1 based on mass analysis data. (B) Western blot analysis of hnRNP A2B1 protein present in 38B9 cells after the indicated treatments. Actin was used as an internal loading control. (C) Whole-cell lysates from 38B9 cells were immunoprecipitated using the anti-MHC class II Ab after the indicated treatments and immunoblotted using the hnRNP A2B1 Ab. The numbers under each gel picture represent the RQ. Representative data from at least 3 independent experiments are presented.
|
hnRNP A2B1 is associated with MAP kinase signaling in 38B9 cells
MAP kinase signaling plays an important role in various cellular processes. We previously demonstrated that ERK/p38 MAP kinases are involved in the proliferation and differentiation of 38B9 cells, especially in MHC class II molecule-mediated negative signaling (11). To confirm the involvement of hnRNP A2B1 in the regulation of MAP kinase signaling, we monitored the influence of siRNA-mediated hnRNP A2B1 inhibition on MAP kinase activation (Fig. 2). As expected, LPS treatment increased the level of hnRNP A2B1 expression in 38B9 B cells by approximately 1.69-fold relative to the control. Additionally, the increased levels of hnRNP A2B1 protein were decreased in hnRNP A2B1 siRNA-treated cells compared with control siRNA-treated cells (both LPS-treated and LPS non-treated) by approximately 0.47- and 0.28-fold, respectively. This suggested that the siRNA targeting hnRNP A2B1 efficiently inhibited hnRNP A2B1 expression. Importantly, knockdown of hnRNP A2B1 influenced phosphorylation of MAP kinases: the levels of p-ERK and p-p38 MAP kinases were increased by LPS by approximately 1.35- and 2.02-fold relative to the control, respectively, while the p-p38 level was decreased by hnRNP A2B1 siRNA by approximately 1.78-fold relative to the control. In contrast, the level of p-JNK MAP kinase was decreased by LPS by approximately 0.17-fold relative to the control but was recovered to 0.96-fold of the control level by hnRNP A2B1 siRNA treatment. Collectively, we confirmed that hnRNP A2B1, which is associated with MHC class II-mediated signaling, is involved in the regulation of MAP kinase signaling in LPS-stimulated 38B9 B cells.
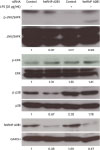 | Figure 2Effect of hnRNP A2B1 on the activation of components of the MAP kinase signaling pathway in 38B9 cells. The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS for 30 min at 37°C. Western blot analyses of the levels of total and p-ERK, p38, and JNK/SAPK in 38B9 cells following the indicated treatments after knocking down hnRNP A2B1 as described in the Materials and Methods section. Actin was used as an internal loading control. The numbers under each gel picture represent the RQ. Representative data from at least 3 independent experiments are presented.
SAPK, stress-activated protein kinase.
|
hnRNP A2B1 is involved in the regulation of NF-κB activation
NF-κB regulates the expression of various cytokine genes and genes involved in the activation, development, and survival of lymphocytes. In addition, LPS is one of the strongest stimulants of NF-κB activation in B cells and is capable of inducing the expression of various cytokines and cell differentiation (16). Because we previously confirmed that anti-MHC class II Ab regulates NF-κB activation in LPS-stimulated 38B9 cells, we monitored the influence of hnRNP A2B1 on NF-κB activation in LPS-stimulated 38B9 cells (Fig. 3A). LPS stimulation of 38B9 cells increased the level of hnRNP A2B1 in the cytoplasm, and the level of hnRNP A2B1 was decreased by siRNA treatment in both the cytoplasm and nucleus. The levels of NF-κB subfamily members c-Rel and p50 in the nucleus were increased by LPS treatment, presumably by their enhanced translocation from the cytoplasm and/or enhanced protein expression by LPS treatment. Additionally, the levels of c-Rel and p50 in both the cytoplasm and nucleus were decreased by hnRNP A2B1 siRNA. Conversely, other NF-κB family members, RelB and p52, showed enhanced translocation into the nuclear compartment after knocking down hnRNP A2B1 using siRNA in LPS-stimulated 38B9 cells. Moreover, the expression of IL-6 and tumor necrosis factor (TNF) was significantly (p<0.01) increased by LPS treatment, and the increased expression of these pro-inflammatory cytokines was inhibited by siRNA-mediated knockdown of hnRNP A2B1 (Fig. 3B), suggesting that hnRNP A2B1 is involved in pro-inflammatory cytokine expression in LPS-stimulated 38B9 cells. Although the activation of NF-κB-related proteins such as c-Rel and p50 was increased following hnRNP A2B1 siRNA treatment, knocking down hnRNP A2B1 did not show any activating effect on the expression of pro-inflammatory cytokines in non-LPS-stimulated 38B9 cells. Collectively, these results suggest that NF-κB activation in LPS-stimulated B cells is negatively regulated by hnRNP A2B1 knockdown, and that NF-κB activation by LPS stimulation in B cells is regulated to recruit hnRNP A2B1 to the cytoplasm.
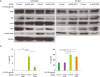 | Figure 3Effect of hnRNP A2B1 siRNA treatment on LPS-mediated NF-κB activation and the expression of pro-inflammatory cytokines in 38B9 cells. (A) The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS for 30 min at 37°C, and proteins in the cytoplasmic and nuclear fractions were analyzed by western blotting. GAPDH was used as an internal control. Representative data from 2 independent experiments are presented. (B) The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS for 12 h at 37°C, and cell culture supernatants were harvested. The levels of IL-6 (left panel) and TNF (right panel) in the culture supernatants were analyzed as described in the Materials and Methods section. Data are presented as means±standard deviation of duplicate determinations, and a representative result from 2 independent experiments is shown.
TNF, tumor necrosis factor.
*p<0.05 and **p<0.01 indicate significant differences between groups.
|
hnRNP A2B1 is involved in Akt signaling in 38B9 cells
The activation of phosphoinositide 3-kinase (PI3K) is required for B cell proliferation and survival. PI3K signaling is also one of the key aspects of B cell differentiation, and its activation promotes Ca2+ mobilization and activation of NF-κB-dependent gene transcription, as well as activation of Akt and mammalian target of rapamycin (mTOR) (4). Because PI3K/Akt activity is involved in B cell differentiation into plasma cells, we analyzed the involvement of hnRNP A2B1 in Akt signaling (Fig. 4). Interestingly, the levels of p-Akt at Thr308 and Ser473 were increased by LPS stimulation by approximately 3.59- and 6.21-fold, respectively, and were inhibited by hnRNP A2B1 siRNA by 1.89- and 3.31-fold, respectively, relative to the control cells. We next analyzed the downstream signaling molecules, including GSK3β and Mcl-1, a myeloid leukemia cell differentiation protein. Similar to p-Akt, the levels of p-GSK3β and Mcl-1 were increased by LPS treatment by approximately 2.01- and 1.68-fold, respectively, and were reduced by hnRNP A2B1 siRNA by 1.39- and 0.51-fold, respectively, relative to the control cells. Collectively, these results suggest that hnRNP A2B1 is involved in cell differentiation via regulation of Akt signaling.
 | Figure 4Effect of hnRNP A2B1 siRNA treatment on components of the Akt signaling pathway in 38B9 cells. The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS at 37°C for 30 min. Western blot analyses of the expression and activation of Akt, GSK3β, and Mcl-1 were performed in cells after the indicated treatments. Actin was used as an internal loading control for western blot analyses. The numbers under each gel picture represent the RQ. Representative data from at least 3 independent experiments are presented.
|
hnRNP A2B1 is involved in B cell maturation
LPS stimulation of B cells via TLR4 induces B cell maturation. 38B9 cells are a pro-B cell line, and the Ig gene is in the rearrangement status of DH-JH alleles but is not induced by VH rearrangement (17). To analyze the influence of hnRNP A2B1 on B cell maturation, we assessed the status of Ig gene rearrangement in LPS-stimulated 38B9 cells after hnRNP A2B1 siRNA treatment (Fig. 5). When we initially measured the level of the germline Igκ light chain gene transcript by RT-PCR, the germline Igκ transcript was not detected in control non-stimulated cells but was detected in LPS-stimulated cells, and the level was reduced in hnRNP A2B1 siRNA-treated cells (Fig. 5A). We confirmed these results by quantitative real-time PCR, and RQ of the germline Igκ transcript was significantly (p<0.05) reduced by approximately 50% in hnRNP A2B1 siRNA-treated cells (Fig. 5B). We next confirmed the expression of surface IgM in 38B9 cells, because germline Igκ expression leads to B cell maturation. As expected, similar to germline Igκ transcript expression, LPS stimulation of 38B9 cells increased the surface IgM-positive cells, and the level of LPS-mediated enhanced surface IgM-positive cells was reduced in hnRNP A2B1 siRNA-treated cells (Fig. 5C). Collectively, these results suggest that hnRNP A2B1 is involved in B cell maturation and differentiation.
 | Figure 5Effect of hnRNP A2B1 silencing on the differentiation of LPS-treated 38B9 cells. The 38B9 cells were transfected with hnRNP A2B1 siRNA. Twenty-four hours after transfection, cells were treated with 25 μg/ml LPS at 37°C for 12 h. Expression of the germline Igκ gene in 38B9 cells after the indicated treatments was determined by (A) RT-PCR and (B) quantitative real-time PCR. In addition, the level of surface IgM expression was determined by flow cytometry. Representative data from at least 3 independent experiments are shown.
*p<0.05 and **p<0.01 indicate significant differences between groups.
|
DISCUSSION
hnRNP A2B1 is an RNA-binding protein and is involved in alternative splicing, cytoplasmic RNA trafficking, and RNA stabilization (18). The hnRNPs are mostly present in the nucleus; however, some hnRNPs, including hnRNP A2B1, seem to shuttle between the nucleus and cytoplasm (19). hnRNP A2B1 also regulates a variety of cellular pathways from cell cycle progression to tissue development in cancer cell lines (20). In this study, we found that hnRNP A2B1 regulated MAP kinase signaling in 38B9 cells (Fig. 2). Specifically, ERK and p38, but not JNK, are closely associated with hnRNP A2B1-associated signaling in LPS-stimulated 38B9 cells. JNK is in an activated state under steady-state conditions and LPS stimulation downregulated JNK. JNK activation appeared to be recovered in LPS-stimulated cells after siRNA-mediated knockdown of hnRNP A2B1, while it was inhibited in non-treated cells after hnRNP A2B1 knockdown (Fig. 2). Therefore, we hypothesize that hnRNP A2B1 functions differently with respect to JNK activation in LPS-stimulated and non-stimulated B cells. Rather, it is tightly associated with the signaling mediated by Akt and the p50 and c-Rel NF-κB subunits, because they showed similar activation patterns as that of JNK in LPS-stimulated and non-stimulated cells (Figs. 3 and 4).
LPS is a potent mitogen for B cell stimulation, promoting extensive proliferation and differentiation of B lymphocytes into Ig-secreting plasma cells (21). B cell differentiation is associated with various factors, including components of the PI3K, Akt, NF-κB, and MAP kinase signaling pathways. The functional and developmental outcomes for B cells, including proliferation, differentiation, and survival, which arise from specific activation signals, reflect the qualitative and quantitative nature of NF-κB responses (16). NF-κB activity is required for 2 distinct stages of B cell development: for viability of pre-B/immature B cell and for providing survival and maturation signals to ensure the proper development of follicular mature B cells. Among the NF-κB components, p50 is involved in pre-BCR-mediated differentiation processes, including Ig light chain expression, while c-Rel expression becomes detectable following pre-B to immature B cell differentiation (22). We determined that either the homo- or heterodimer of p50/c-Rel was translocated to the nucleus by LPS treatment, and that LPS-mediated NF-κB activation was inhibited by knocking down hnRNP A2B1 (Fig. 2). Similarly, Igκ light chain and surface IgM expression was decreased by hnRNP A2B1 knockdown in LPS-stimulated 39B9 cells (Fig. 5). Consequently, we believe that p50/c-Rel activation was impaired by hnRNP A2B1 knockdown, resulting in suppression of 38B9 B cell differentiation. (Fig. 6)
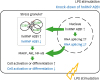 | Figure 6Schematic diagram outlining possible regulatory mechanism of cellular responses by hnRNP A2B1 in LPS-stimulated 38B9 cells. LPS stimulation of 38B9 cells increases the level of hnRNP A2B1 protein and may accumulate in stress granule in cytoplasm, which, in turn, activates cellular signaling pathways to activate 38B9 cells. These signaling pathways are suppressed by knocking-down of hnRNP A2B1.
|
The Akt pathway is highly associated with B cell differentiation process. Although p-PDK1 was consistently upregulated in 38B9 cells and was reduced by LPS treatment, Akt phosphorylation was increased by LPS treatment (data not shown). This phenomenon of disharmony between Akt and PDK1 was similarly observed in some cancer cells, and several Akt-independent mechanisms used by PI3K to promote cancer growth have been described (23). These PDK1-independent Akt activation mechanisms include activation by Ack I, IKKe, TBK1, Src, or PTK6 in various cancer cells. Given that the 38B9 cell line is an A-MuLV-transformed cell line, it is plausible that 38B9 cells always exhibit upregulation of PDK1 and induce PDK1-independent Akt activation. PDK1-independent phosphorylation of Akt induces phosphorylation of GSK3β, and GSK3β phosphorylation inhibits self-kinase activity. Importantly, GSK3β phosphorylation prevents the phosphorylation-dependent degradation of Mcl-1, which is required for survival throughout B cell development (24). Akt phosphorylation is regulated by hnRNP A2B1 in LPS-stimulated 38B9 cells, and the decrease in Akt phosphorylation induced by hnRNP A2B1 siRNA led to inhibition of GSK3β phosphorylation, thereby decreasing Mcl-1 expression in LPS-stimulated 38B9 cells (Fig. 4). Thus, we speculate that hnRNP A2B1 regulates B cell differentiation via the Akt and NF-κB signaling pathways.
The cellular function of hnRNP A2B1 is well-recognized in the nucleus as an important regulator of transcripts containing an A2 response element or A2 response element-like sequences, and hnRNP A2B1 controls the ERK MAP kinase pathway via A-Raf splicing (25). Consequently, it is believed that hnRNP A2B1 regulates cell signaling through controlling RNA splicing in nucleus. However, we found that cytoplasmic accumulation of hnRNP A2B1 in LPS-stimulatedcells (Fig. 3), and were interested in the function of hnRNP A2B1 in cytoplasm because the role of hnRNP A2B1 in the cytoplasm remains poorly understood. It was recently reported that hnRNP A2B1 and hnRNP A1 contain a prion-like domain enriched with uncharged polar amino acids and glycine and are recruited to stress granules under stress conditions by arsenite treatment (26). We also observed cytoplasmic hnRNP A2B1 and G3BP1, which is a stress granule marker, together in MHC class II-associated proteins, which was confirmed by immunoprecipitation and confocal laser scanning microscopy (data not shown). Moreover, osmotic stress activates the MEK3/6-p38 kinase pathway via Mnk kinase, which in turn, evokes the cytoplasmic accumulation of hnRNP A2B1 (27). Given that we also found a positive correlation between hnRNP A2B1 translocation to the cytoplasm and p38 MAP kinase phosphorylation, we believe that hnRNP A2B1 may function differently in the cytoplasm and nucleus.
Collectively, we identified hnRNP A2B1 as an MHC class II-associated protein using a proteomics strategy and demonstrated that hnRNP A2B1 is associated with B cell differentiation by regulating MAP kinase signaling, NF-κB activation, and the Akt signaling pathway. In addition, we speculate that hnRNP A2B1 exerts this specific function in the signaling pathways because it is recruited to the cytoplasm. Although we did not directly confirm the role of cytoplasmic hnRNP A2B1, we hypothesize that hnRNP A2B1 regulates these cellular pathways by accumulating stress granules. We also believe that MHC class II-mediated molecules, including hnRNP A2B1, regulate B cell activation and may be associated with the recruitment of stress granules.
 XML Download
XML Download