

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
E. coli
Escherichia coli
IFN
interferon
IPTG
isopropyl β-D-1-thiogalactopyranoside
IRF
interferon regulatory factor
ISG
interferon-stimulated gene
LPS
Lipopolysaccharide
MDBK
Madin-Darby bovine kidney epithelial
MDCK
Madin-Darby canine kidney epithelial-like
OAS
oligoadenylate synthetase
JAK
Janus kinase
STAT
signal transducers and activators of transcription
VSV
vesicular stomatitis virus
WISH
Wistar Institute Susan Hayflick
INTRODUCTION
Interferon (IFN) subclasses exist in vertebrates that have antiviral properties and cell-mediated immunity varying with cell types, animal species, and viruses. The type 1 IFN constitutes a large multigene family that includes IFNα, IFNβ, IFNτ, and IFNω. It is known that these 4 members of type 1 IFNs activate common cell surface receptors, IFNaR1 signal transducing chain and IFNaR2 ligand binding chain, respectively (123). Unlike vitamin and some hormone, cytokines need cell surface receptors to activate downstream signaling in order to induce immune responses against different types of pathogen such as virus, bacteria, fungus, and parasite (4567).
IFNγ is only type 2 IFN that binds to distinct receptors and locates a separate chromosomal locus. Initially, IFNγ was considered as a molecule produced from CD4+ T helper cell type 1 lymphocytes, CD8+ cytotoxic lymphocytes, and natural killer (NK) cells exclusively (8910). IFNγ from professional antigen presenting cells (APCs) such as monocyte, macrophage and dendritic cells acting locally has an important function in cell self-activation and activation of nearby cells (1112). T lymphocytes are the major source of IFNγ in the acquired immunity although IFNγ is produced by NK cells. Possibly professional APCs are likely to be a part of function in early host defense against infections (1113).
IFNλs are classified as type 3 IFNs (known as IL-28 and IL-29) and they are cytokines with IFN-like activities. Several types of viruses induce IFNλ1 and IFNλ2/3 in similar patterns. The IFNλs were characterized as interferon-stimulated genes (ISGs) unlike direct induction of IFNα/β by viral infection, thus identifying type 3 IFNs. In vitro assays revealed that IFNλs have detectable antiviral function against encephalomyocarditis virus, but they have limited activities against herpes simplex virus type 2 unlike potent inhibition of both viruses by IFNα (14).
The cytosolic part of receptor is associated with Janus kinase (JAK) activating after type 1 IFN binding to IFNaR1/2 that subsequently phosphorylates downstream signaling molecule named signal transducers and activators of transcription (STAT). The dimerization of STAT interacts with interferon regulatory factor (IRF)-9 to form a trimeric interferon-stimulated gene factor-3 (ISGF-3) complex translocating into the nucleus where it binds an interferon-stimulated regulatory element (ISRE) resulting in the induction of IRF-1 gene (15). The induction of IRF-1 drives the expression of IRF-2, which interacts with other regulatory elements to control the expression of type 1 IFN responsive genes (1617).
IFNτ also belongs to type 1 IFN that present only in ruminants. Among type 1 IFNs, IFNτ amino acid sequence exhibits the highest homology with IFNω, but also shares a limited identity with IFNα/β. The amino acid sequence of IFNτ shares 50% and 25% identity with IFNα/β, respectively (18). IFNτ binds to the common type 1 IFN receptors, a hetero dimeric form of IFNaR1 (19) and IFNaR2 (2). However, IFNτ is produced mainly from trophoblasts of ruminant conceptuses during the blastocyst stage when the elongated trophoblast attaches to the uterine wall. The secretion of IFNτ prevents the destruction of the corpus luteum and helps in maternal recognition of pregnancy. In ruminant reproduction, IFNτ constitutes an actual pregnancy signal (2021222324). In the pregnant period, IFNs produced by trophoblast are detected in other mammals but antiviral activity is almost non-detectable (25).
Type 1 IFNs induced antiviral activity via inhibiting viral gene transcription. The common ISGs associated with antiviral activities are 1) double stranded RNA-dependent protein kinase (PKR) suppressing translation initiation through the phosphorylation of protein synthesis initiation factor eIF-2α, 2) 2′,5′-oligoadenylate synthetase (OAS)-1 family and RNase L nuclease, which mediate RNA degradation, and 3) Mx dynamin-like GTPase (Mx) family targeting viral nucleocapsids resulted in inhibition of RNA synthesis (26272829). In the present study, we cloned porcine IFNα8 from domestic pig (Sus scrofa domestica), a main strain in Korea, and compared its antiviral activities to human IFNα2 using different species of cell lines.
MATERIALS AND METHODS
Cells and reagents
Human amnion (fibroblast) Wistar Institute Susan Hayflick (WISH) cells, Madin-Darby bovine kidney (MDBK) epithelial cells, Madin-Darby canine kidney (MDCK) epithelial-like cell, and vesicular stomatitis virus (VSV; Indiana strain) were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). WISH, MDBK, and MDCK cells were cultured in a medium containing 1% penicillin-streptomycin (Life Technologies, Grand Island, NY, USA), and 10% fetal bovine serum (Hyclone, Logan, UT, USA) according to the ATCC's instructions. Human recombinant IFNα2 protein was obtained from LG biotech (Seoul, Korea). The IgG-horseradish peroxidase (HRP) conjugated secondary antibodies and anti-actin primary antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
RNA extraction and RT-PCR for TA cloning
Total RNA was isolated from whole blood of domestic pig (Sus scrofa domestica) with the QIAamp RNA Blood Mini Kit (Qiagen, Valencia, CA, USA). Total RNA (2 µg) was reverse-transcribed with 0.5 µl of Superscript II reverse transcriptase (Invitrogen, Carlsbad, CA, USA) in 20 µl reaction volume as described previously (30). The 2 µl of cDNA was propagated to perform PCR of porcine IFNα8 (accession No. ACV42397) with sense primer: 5′-AGCATCTGCAAGGTTCCCAA-3′; reverse primer: 5′-CAGGTGTCTGTCACTCCTTC-3′. PCR mixture was denatured at 94°C for 20 s, annealed at 59°C for 40 s, elongated at 72°C for 1 min, and reacted 30 cycles. The PCR products were resolved by 1%-agarose gel electrophoresis. The PCR product was visualized under ultra-violet (UV) trans-illuminator.
Construction of IFNα8 expression vector
The RT-PCR product of porcine IFNα8 was ligated into T&A cloning vector (RBC Bioscience, Xindian City, Taiwan) and the insert of cDNA was verified with DNA sequencing analysis (COSMO Genetech, Seoul, Korea). The region of mature IFNα8 without signal sequence was amplified by PCR method with primers containing EcoRI recognition site at 5′ end and XbaI recognition site at 3′ end (sense primer: 5′-ATATGAATTCTGCGACCTGCCTCAGA-3′; reverse primer: 5′-TATATCTAGAT CACTCCTTCCTCCTG-3′). The PCR product was trimmed with EcoRI and XbaI, and ligated into pProEx/HTa expression vector (Invitrogen). The sequence of mature porcine pProEx/HTa-IFNα8 was verified by DNA sequencing prior to using for protein expression.
Expression and purification of recombinant proteins
The mature porcine pProEx/HTa-IFNα8 Escherichia coli (E. coli) expression vector was transferred into BL21/Codon and BL21/Rosetta (Promega, Fitchburg, WI, USA) because the yield of recombinant porcine IFNα8 in DH5α was not good enough (data not shown). The clones of porcine IFNα8 BL21/Codon plus and BL21/Rosetta were cultured in 1 ml volume of Luria-Bertani (LB) broth. The pilot experiment examined the level of recombinant protein in insoluble pellet or soluble supernatant by western blotting as described (31).
Porcine IFNα8 was expressed in BL21/Codon plus since this host strain produced the great amount of recombinant protein. The cells were cultured at 37°C. When the OD of cultured broth at 600 nm reached 0.6 OD, isopropyl β-D-1-thiogalactopyranoside (IPTG) was added to induce the target protein expression for 3 h at 37°C. After 3 h, the cells were collected by centrifugation at 8,000 rpm for 15 min at 4°C, resuspended in basic buffer (8 M Urea, 20 mM Tris-Hcl, pH 9.0), and subjected to ultrasonication. After 3 h incubation at 37°C, the supernatant was collected by centrifugation at 10,000 rpm for 20 min at 4°C and followed by sonication (pulse 30 s and interval 30 s for 2 min). It was then centrifuged at 10,000 rpm for 10 min at 20°C, when the supernatant was collected for purification. The recombinant porcine IFNα8 protein was purified with a Talon affinity column (Invitrogen) using his6-tag at N-terminus of the recombinant protein. Before loading the collected supernatant, the basic buffer was loaded onto Talon column for washing and equilibrium. The supernatant was loaded onto the column twice, and 0.1% Triton X 114 in basic buffer was loaded to remove lipopolysaccharide (LPS) (32). After washing with basic buffer, the protein was eluted with elution buffer (8 M Urea, 20 mM Tris-Hcl, pH 9.0, 150 mM imidazole). Four fractions (1 ml per fraction) were collected. After sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), the protein bands were visualized by staining with Coomassie brilliant blue. After Talon affinity-purified porcine IFNα8 dialyzed in 20 mM Tris-Hcl, pH 9.0 then applied to anion exchange chromatography (GE Healthcare, Marlborough, MA, USA). The recombinant protein was analyzed by western blot using mouse monoclonal antibody against his6-tag (R&D system, Minneapolis, MN, USA) as described (4).
Cytopathic effect of antiviral assay
To determine the antiviral activity of recombinant porcine IFNα8, vesicular stomatitis virus (VSV) inhibition assay was performed with canine MDCK, bovine MDBK and human WISH cells. These cells were cultured in 96-well plates until the cells reached monolayer status. Then the cells were washed and bovine porcine IFNα8 proteins were added to the plate. Human IFNα2 was used as a positive control. Porcine IFNα8 proteins and human IFNα2 were used to treat the cell by a 2-fold serial dilution. The treated cells were incubated for 6 h and then VSV was added for infection. After 22 h of VSV infection, the media was removed and the cells were stained with crystal violet solution.
RT-PCR
MDBK, WISH, and MDCK cells (1×106 cells/well in 6-well plates; TPP Techno Plastic Products AG, Trasadingen, Switzerland) were treated with human porcine IFNα8 (20 ng/ml) at different time points (3 or 6 h) and then harvested for RNA extraction. Total RNA was isolated with Tri reagent (Sigma-Aldrich, St. Louis, MO, USA) according to manufactures manual. Total RNA (2 µg) samples were used for RT-PCR. 2 µl of cDNA was used to perform PCR of canine MX dynamin-like GTPase 1 (Mx-1, accession No. NM_001003134) with sense primer: 5′-TGGAGGCTCTGTCAGGAGTT-3′; reverse primer: 5′-TTGCCTTC AGTCCCTCTGTC-3′, canine OAS-1 (accession No. AY863104) with sense primer: 5′-ATCTCCTGCCAGACACA CAG-3′; reverse primer: 5′-GTGAAGCAGGTGGAGAACTC-3′, canine β-actin (accession No. NM_001195845) with sense primer: 5′-ACCAACTGGGACGACATGGA-3′; reverse primer: 5′-GCCAATAGTGATGACCTGCC-3′. The rest of primers for Mx-1, OAS-1, and β-actin have been described in previous study (33).
RESULTS
Constitutive expression of IFNα in porcine peripheral whole blood cells
First, we examined the regulation of porcine IFNα8 with fresh peripheral whole blood cells from Korean domestic pig (Sus scrofa domestica). The whole blood cells were treated with different stimuli as indicated on the top. Interestingly, constitutive expression of porcine IFNα8 was detected in unstimulated control and there was no significant induction of the transcript (Fig. 1). The control of β-actin exhibited at bottom panel to show that the difference is not due to varying amount of transcript. We used the RT-PCR product of the untreated control transcript for T&A cloning and its DNA sequence was verified as described in Materials and Methods section. As shown in Fig. 2, porcine IFN open reading frame (ORF) contains 570 base pair including first codon for “Met” residue and the last stop codon “TGA”. The 23 amino acid sequence of hydrophobic signal peptide was highlighted with green color.
 | Figure 1The regulation of porcine IFNα8 transcript. RT-PCR was performed with the whole blood RNA from Korean domestic pig (Sus scrofa domestica) under stimulation of LPS, polyinosinic-polycytidylic acid (Poly I:C), and VSV at time point of 3 h. The control β-actin showed no difference in transcript. The data represents one of 4 independent experiments.
Ctl, control; LPS, lipopolysaccharide.
|
 | Figure 2Translated amino acid sequence of porcine IFNα. The nucleotide sequence was used to convert into amino acid sequence. The hydrophobic signal sequence of 69 base pairs nucleotide sequences were highlighted by green color with underlined 23 amino acid residues. This sequence was deposited in databank and obtained accession number as “KX275310”.
|
Amino acid sequence of IFNα8 from Korean domestic pig
The obtained amino acid sequence of porcine IFNα was aligned with porcine IFNα8 in Fig. 3A. The amino acid sequence of T&A cloned the porcine IFNα8 from Korean domestic pig shared 98.4% identity with the known porcine IFNα8 (accession No. ACV42397) as shown in Fig. 3. Three amino acid residues are different from the known porcine IFNα8 and the different residues are highlighted by yellow. The newly obtained Korean porcine IFNα8 was registered in databank as accession number (KX275310). The region of mature protein without signal sequence (Met/1-Gly/23) was subcloned into pProEx/HTa vector for E. coli expression. Actually, 2 amino acid residues of Korean porcine mature IFNα8 are different from that of the known IFNα8 in databank.
 | Figure 3Comparison of porcine IFNα8 to the known porcine IFNα. The ORF of porcine IFNα8 mRNA from Korean domestic pig was analyzed and newly identified as porcine IFNα8 (accession No. KX275310). The amino acid sequence was deduced by newly obtained the DNA sequence of porcine IFNα8 and the different amino acid residue of Korean porcine IFNα8 compared to porcine IFNα8 (accession No. ACV42397). Three distinct amino acid residues are highlighted with yellow color.
ORF, open reading frame.
|
Expression of recombinant porcine IFNα8 protein
In order to compare the expression efficiency of recombinant IFNα8 protein in E. coli, the expression vectors were transformed into E. coli BL21/Codon and Rosetta strain. The production of recombinant proteins was induced by adding IPTG and the result was confirmed by western blot analysis by using anti-his6-tag antibody (Fig. 4A). The anti-his6-tag antibody recognized the IPTG-induced 25 kDa band as indicated by arrow. A large-scale expression was processed with BL21/Codon plus strain and the production of recombinant porcine IFNα8 proteins was present in an insoluble fraction (not shown).
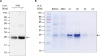 | Figure 4The expression and purification of recombinant porcine IFNα8 with Talon affinity chromatography. The recombinant porcine IFNα8 mRNA from Korean domestic pig was expressed in E. coli and purified by a mini-Talon metal affinity column. (A) The IPTG-induced recombinant porcine IFNα8 detected with mouse anti-his6-tag monoclonal antibody. The arrow indicated porcine IFNα8 induced in the presence of IPTG for 3 h. (B) The a mini-Talon metal affinity chromatography purified recombinant porcine IFNα8 was visualized by Coomassie blue staining. The data represents one of 5 independent experiments.
|
The first step of purification was performed by using a mini-Talon affinity column. The eluted fractions from the Talon affinity column were analyzed by Coomassie blue staining (Fig. 4B). The molecular weight of porcine IFNα8 appeared to be approximately 25 kDa band (see the arrow in Fig. 4B). The purified recombinant IFNα8 protein by the Talon affinity column was further purified by anion exchange chromatography after overnight dialysis against Tris-Hcl (20 mM, pH 9.0). The elution peak of recombinant porcine IFNα8 protein was observed mainly in bound fractions (Fig. 5A). The anion exchange chromatography fractions were visualized by silver staining to confirm the purity of the recombinant porcine IFNα8 protein (Fig. 5B). The anion exchange chromatography purified recombinant porcine IFNα8 protein appeared to be approximately 25 kDa and faint 38 kDa band in silver staining. We confirmed 38 kDa band is porcine IFNα8 by western blot (data not shown). The fractions were pooled as unbound fractions for quantification comparing to bovine serum albumin (BSA) in Fig. 5C.
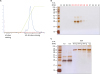 | Figure 5The purification of porcine IFNα8 by anion exchange chromatography. (A) UV absorbance at 280 nm was recorded during the porcine IFNα8 the second step purification process with anion chromatography. (B) The purified recombinant porcine IFNα8 fractions were visualized by silver staining. (C) The bound fractions of recombinant porcine IFNα8 were pooled for quantification to compare with BSA in silver staining. The data represents one of 5 independent experiments.
BSA, bovine serum albumin; UV, ultra-violet.
|
Antiviral activity of porcine IFNα8 against VSV
The antiviral assay was first performed with human WISH cells by using both porcine IFNα8 and the control human IFNα2. As shown in Fig. 6A, human IFNα2 protected the cells from VSV infection while porcine IFNα8 failed to protect the cells. Next, we used bovine MDBK cells for porcine IFNα8 antiviral assay and both porcine IFNα8 and human IFNα2 protected MDBK cells (Fig. 6B). We further examined canine MDCK cells and human IFNα2 and porcine IFNα8 failed to protect MCBK cells from VSV infection (Fig. 6C).
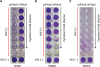 | Figure 6Recombinant porcine IFNα8 reduces the cytopathic effect caused by VSV. (A) Human epithelial WISH, (B) bovine MDBK, (C) canine MDCK cells were treated with recombinant porcine IFNα8 including positive control human IFNα2 then infected with VSV to analyze the reduction of cytopathic effect. The plate was visualized with crystal violet staining. The concentration of porcine IFNα8 (20 ng/ml) and human IFNα2 (2 ng/ml) were added by the 2-fold serial dilution. For control, the wells at bottom row were non-infected cells; the rows above non-infected wells were only VSV infected cells which shows the complete cytopathic effect to VSV challenge. The data represents one of 5 independent experiments.
hIFN, human interferon; pIFN, porcine interferon.
|
The antiviral activity of porcine IFNα8 via Mx-1 and OAS-1
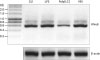
To investigate the antiviral mechanism of porcine IFNα8, the induction of ISGs was tested with RT-PCR. The levels of Mx-1 and OAS-1 were increased in bovine MDBK cells treated with porcine IFNα8 at 3 and 6 h (Fig. 7A) however the induction of antiviral genes in human WISH cells increased with only human IFNα2 (Fig. 7B). Interestingly, the high constitutive expression of Mx-1 and OAS-1 in canine MCBK was observed in the absence of IFNs stimulation (Fig. 7C). The control β-actin transcript showed that the induction of ISGs is not due to experimental variation (Fig. 7).
 | Figure 7Recombinant porcine IFNα8 induces antiviral genes. (A) Bovine MDBK, (B) human WISH, and (C) canine MDCK cells were treated with human porcine IFNα8 (20 ng/ml) and human IFNα2 (2 ng/ml) at indicated time points (3 and 6 h). Non-treated cells were prepared as negative control. After treatment of IFNs, total RNA was extracted for RT-PCR of Mx-1, OAS-1, and β-actin for normalization. The data represents one of 3 independent experiments.
Ctl, control; hIFN, human interferon; pIFN, porcine interferon.
|
DISCUSSION
In the present study, we cloned porcine IFNα8 from domestic pig (Sus scrofa domestica), main strain of pig in Korea. The recombinant porcine IFNα8 protein was expressed in E. coli and examined for its antiviral activity with different species of cell line. Interestingly, the amino acid sequence of porcine IFNα8 is different from that of the known porcine IFNα8 in databank (accession No. ACV42397). The amino acid sequence of porcine IFNα8 shares 98.4% identity with the known porcine IFNα8. We deposited newly cloned porcine IFNα8 from Korean domestic pig (Sus scrofa domestica) in gene bank and obtained its accession number (KX275310).
Porcine recombinant IFNα8 was examined for its biological activity using a common antiviral assay. The antiviral activity of porcine IFNα8 was weaker in its cytopathic effect compared to human IFNα2 in bovine MDBK cells. The antiviral activity of porcine IFNα8 was obtained only with MDBK cells however human IFNα2 exhibited antiviral activity in both human WISH and bovine MDBK cells (Fig. 6). In addition, both porcine IFNα8 and human IFNα2 failed to show antiviral activity in canine MDCK cells (Fig. 6). In addition, the results of RT-PCR in Fig. 7 corresponded to the antiviral activity of IFNs in Fig. 6. These data suggested the species specificity with different IFNs.
It has been reported that human type 1 IFNs is superior in their activities in different species whereas mouse type 1 IFN inferior to other species such as human cells. This species-specific activity provided Uzé et al. (19), to isolate IFNaR1 more than 20 years ago. However, there was no difference of activity of porcine IFNα8 and human IFNα2 in canine MDCK cell and both IFNs failed to show the antiviral activities in this cell line (Fig. 6C). This result may be explained by RT-PCR data that the high constitutive expression of antiviral genes without IFN stimulation (Fig. 7C).
Although IFNα, IFNβ, IFNω, and IFNτ bind to common type 1 receptors, IFNaR1 and IFNaR2, their bindings and biological activities clearly exhibit species-, tissue-, and cell- specific differences probably due to 3-dimensional conformation differences among type 1 IFN ligands (18). Thus, the result indicated that the low activity of porcine IFNα8 in MDBK cells could be due to the difference of amino acid sequence between porcine IFNα8 and human IFNα2. However, we cannot exclude that there are additional unknown receptor components contributing for the species-specific antiviral activity of porcine IFNα8. The biological activity of porcine IFNα8 would be optimized in cells of their own strain bearing high-affinity receptors. It is necessary to investigate whether there is an additional receptor like IL-1 family cytokine ligands and receptors possessing 11 members of ligands and receptors (5) while 4 distinct ligands of type 1 IFN has only 2 receptors components.
Type 1 IFNs induce the synthesis of antiviral factors involving OAS-1 and Mx-1 genes via activation of the cellular JAK-STAT signaling pathway (234). The IFN-inducible OAS leading to the degradation of RNA requires 2 enzymes, OAS, and RNase L (35). Three isoforms of OAS, designated as OAS-1, OAS-2, and OAS-3, have been identified in human cells by immunoblotting and by characterization of cDNA and genomic clone analysis (3637), however it is known that the oligomerization of OAS-1 and OAS-2 appears necessary for enzymatic activity (38394041). RNase L, a latent endoribonuclease, becomes active by binding 2–5 A oligonucleotides and digests the unusual RNAs. We found that porcine IFNα8 and human IFNα2 increased the synthesis of the cellular antiviral factors, such as OAS and Mx-1 proteins (Fig. 7). These results suggest that porcine IFNα8 induces the antiviral activity through similar pathways like other type 1 IFNs.
This study describes the cloning and characterization of porcine IFNα8 from domestic pig in Korea. Unexpectedly, the amino acid sequence of the porcine IFNα8 differs by about 2% from that of the known porcine IFNα8 in databank. We expect that the recombinant porcine IFNα8 would prevent different virus infection with lower cytotoxicity than other existing therapies. A large quantity of biologically active recombinant IFNα8 protein can be also used to develop protein drug for preventing early viral infection in industrial baby pig.
 XML Download
XML Download