

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Confronted with various kinds of pathogens, serosal cavities harbor distinctive types of lymphocytes including innate-like B-1a cells as well as innate immune cells, such as mast cells, macrophages, and natural killer (NK) cells (12). Recently, we reported the abundance of innate-like CD4+ T cells in the peritoneal cavity (PEC), which characteristically expressed a high level of integrin α4 (CD49d) and rapidly secreted Th1 cytokines upon stimulation similarly to memory T cells (3). The innate-like integrin α4highCD4+ T cells developed very early before the age of 3 days and provided help for B-1a cells. These peritoneal innate-like B-1a and integrin α4highCD4+ T cells are presumed to be responsible for the immediate response to pathogenic invasion into the PEC that results from uncontrolled gastrointestinal infection. The pleural cavity (PLC) similarly harbors innate-like lymphocytes including B-1a cells that had previously described as innate response activator B cells (4), but the characteristics of PLC T cells are not well investigated.
Integrin α4 is an integrin chain that can combine with either of 2 β chains, β1 (CD29) or β7, to form integrin α4β1 (VLA-4) or integrin α4β7 (LPAM-1) heterodimers, respectively (5). Both integrin heterodimers are not highly expressed on naive lymphocytes, but highly on specific types of memory lymphocytes (6). Integrin α4β1 or α4β7 directs transendothelial migration into distinct anatomical sites; α4β1 through binding to vascular cell adhesion protein 1 (VCAM-1) for the entry into inflammatory sites and α4β7 through binding to mucosal vascular addressin cell adhesion molecule-1 (MAdCAM-1) for recirculation to sites of intestinal inflammation and intestinal secondary lymphoid tissues (7). In human memory T cells, the expression of α4β1 is reciprocally correlated to the expression of α4β7 (8). Therefore, the fine regulation of the integrin β chain expression is important for the preferential recruitment of memory lymphocytes and intestinal pathology (910).
On the other hand, integrin β1 can combine with at least 12 α integrin chains to form different kinds of integrin combinations that bind to extracellular matrix proteins, such as α1β1 (VLA-1), α2β1 (VLA-2), α3β1 (VLA-3), α4β1 (VLA-4), α5β1 (VLA-5), and α6β1 (VLA-6) (11). Since the adhesive activity of these β1 integrins is enhanced by T cell receptor (TCR)-mediated signaling, the expression of β1 integrins is important for the function of memory/effector T cells that express a higher level of integrin β1 than naïve T cells (12). What determines the kind of integrin α chain expressed in combination with integrin β1 is not well understood, but the expression of different β1 integrin complexes regulates their adhesive and migratory behavior of memory T cells (131415).
In this study, we systemically analyzed the expression of various kinds of integrin chains in the serosal integrin α4highCD4+ T cells. We observed that most of them preferentially expressed the α4β1 and α6β1 integrins, but a minor population of integrin α4highCD4+ T cells expressed a high level of α4β7 integrin. We further reanalyzed the functional differences of 2 kinds of serosal integrin α4highCD4+ T cells.
MATERIALS AND METHODS
Mice
C57BL/6 mice were purchased from Orient Bio (Sungnam, Korea). Cd1d−/− mice were kindly provided by L. Van Kaer (Vanderbilt University School of Medicine). This study was approved by the Institutional Animal Care and Use Committee of Sungkyunkwan University School of Medicine. All procedures were performed in a pathogen-free facility, according to institutional guidelines.
Cell preparation and flow cytometric analysis
Peritoneal and pleural cells were isolated by flushing the serosal cavities with PBS. Cells were stained on ice for 30 min with the appropriate combinations of fluorochrome-conjugated Abs in FACS buffer (5% bovine calf serum [BCS] and 0.05% sodium azide in PBS). Following fluorochrome-labeled monoclonal antibodies were used: CXCR6 (221002), α3 (polyclonal), α7 (334908) from BD Biosciences (San Jose, CA, USA); CD4 (GK1.5), CD62L (MEL-14), CD44 (IM7), CXCR3 (CXCR3-173), CCR4 (2G12), CCR5 (HM-CCR5), PD-1 (29F.1A12), SLAM (TC15-12F12.2), CD122 (5H4), CD127 (A7R34), α1 (HMa1), α2 (DX5), α5 (5H10-27, MFR5), α6 (GoH3), β1 (HM81-1), β7 (FIB504), α4β7 integrin (DATK32), interferon (IFN)-γ (XMG1.2), IL-10 (JES5-16E3), TGF-β (TW7-20B9), tumor necrosis factor (TNF)-α (MP6-XT22) from BioLegend (San Diego, CA, USA); and CXCR4 (2B11), CXCR5 (SPRCL5), CCR6 (Slrx6), CCR7 (4B12), ICOS (7E.17G9), α4 (R1-2), IL-4 (11B11) from eBioscience (San Diego, CA, USA). After washing with FACS buffer, the stained cells were analyzed on a FACSCanto II system (BD Biosciences). Data were analyzed using FlowJo software (Tree Star, San Carlos, CA, USA).
Intracellular staining for cytokines
Cells were suspended in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS), 25 mM sodium bicarbonate, 2 mM glutamine, 50 U/ml penicillin, 50 mg/ml streptomycin, and 10 mM HEPES (all from Invitrogen Life Technologies, Carlsbad, CA, USA) and then stimulated with 50 ng/ml PMA (Sigma-Aldrich, St. Louis, MO, USA) and 1.5 mM ionomycin (Sigma-Aldrich) for 4 h. Brefeldin A (Sigma-Aldrich) was added to 10 µg/ml during the last 3 h of stimulation. Cells were stained with anti-CD4, β1, β7, and anti-α4 Abs, fixed with 2% paraformaldehyde in PBS, permeabilized with 0.1% BSA/0.05% Triton X-100 in PBS, and stained with Abs against IFN-γ, IL-2, IL-4, IL-5, IL-10, IL-13, TGF-β, or TNF-α.
Statistical analysis
Student's t-test (unpaired) and 1-way or 2-way ANOVA tests were used to assess the statistical significance of differences between groups. The p-values <0.05 were considered to be statistically significant for all tests. Histograms were plotted using GraphPad Prism 4.0 (GraphPad Software, San Diego, CA, USA).
RESULTS
PLC contains integrin α4highCD4+ T cells with a memory phenotype similar to that of peritoneal integrin α4highCD4+ T cells
Since B-1a cells are abundant in PLC as well as PEC, we first investigated whether the integrin α4highCD4+ T cells are present in the PLC. The integrin α4highCD4+ T cells were also abundant in the PLC although their proportion among CD4+ T cells was smaller in the PLC than in the PEC (Fig. 1). As expected, the pleural integrin α4highCD4+ T cells cavity also showed the memory and pro-inflammatory phenotypes found in the peritoneal integrin α4highCD4+ T cells, which include a high expression of CD44, CXCR3, PD-1, ICOS, SLAM, CD122, and CD127, a low expression of CD62L, and a rapid secretion of IFN-γ upon stimulation (data not shown). These cells were not natural killer T (NKT) cells as they were present in Cd1d−/− mice.
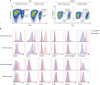 | Figure 1Expression of activation-related cell surface molecules in peritoneal and pleural integrin α4highCD4+ T cells. (A) PEC and PLC cells obtained from 8-week-old C57BL/6 WT or Cd1d
−/− mice were examined for the expression of CD4 and integrin α4 (CD49d). (B) Expression of given cell surface proteins in gated peritoneal and pleural integrin α4highCD4+ T cells (red line, CD49dhigh) or CD49dlow T cells (blue line, CD49dlow). Data are representative of 12 separate experiments.
WT, wild type.
|
The serosal integrin α4highCD4+ T cells are divided into 2 populations based on the expression of α6β1 or α4β7 integrins
We next investigated the expression of integrin β1 or β7 chains in the serosal integrin α4highCD4+ T cells since the integrin α4 is known to heterodimerize with integrin β1 or β7. When we checked the expression of integrin β1 or β7 chains in serosal CD4+ T cells, the memory phenotype CD4+ T cells were recognized as integrin β1+CD4+ T cells (Fig. 2A). The integrin β1−CD4+ T cells were not found in the gated integrin α4highCD4+ T cells, which indicates that the integrin α4β1 is a principal integrin complex in the serosal integrin α4highCD4+ T cells. Notably, a small population of integrin α4highβ1+β7+CD4+ T cells was also observed. The level of the integrin β1 expression is higher in the integrin α4highβ1+β7−CD4+ T cells than in α4highβ1+β7+CD4+ T cells. Therefore, we divided the integrin α4highCD4+ T cells into α4highβ1highCD4+ and α4highβ1+β7+CD4+ T cells. We presumed that some fraction of integrin β1 in the integrin α4highβ1highCD4+ T cells might associate with some other integrin α chain(s) to form other β1 integrin complexes.
 | Figure 2Serosal integrin α4highCD4+ T cells are divided into integrin α4highβ1highCD4+ T and α4highβ1+β7+CD4+ T cells. (A) PEC and PLC CD4+ T cells or gated integrin α4highCD4+ T cells obtained from 8-week-old C57BL/6 wild type mice were shown for the expression of integrin β1 and β7. (B) Illustration for integrin combinations of integrin α4, β1, and β7 chains. (C) Cell surface expression of various integrin α chains in integrin α4highβ1highCD4+ T cells (red line, α4highβ1high), integrin α4highβ1+β7+CD4+ T cells (blue line, α4highβ1+β7+), and integrin α4lowCD4+ T cells (green line, α4low). Data are representative of at least 3 separate experiments.
|
In contrast to integrin α4 that associate with only 2 kinds of integrin β chains, the integrin β1 chain can associate with many integrin α chains (Fig. 2B). To find additional integrin β1 combination(s) in the integrin α4highβ1highCD4+ T cells, we investigated the expression of individual integrin α chains in the 2 integrin α4highCD4+ T cell populations as well as integrin α4lowCD4+ T cells (Fig. 2C). Notably, the high expression of integrin α6 is prominent only in the integrin α4highβ1highCD4+ T cells, but not in the other populations. This result suggests that α6β1 (VLA-6) is another inflammatory integrin chain expressed on the serosal integrin α4highβ1highCD4+ T cells.
The serosal integrin α4highβ1highCD4+ and α4highβ1+β7+CD4+ T cells are distinct memory phenotype T cells
We addressed whether the integrin α4highβ1highCD4+ and α4highβ1+β7+CD4+ T cells were functionally distinct or not. We compared the expression of activation-related molecules in the 2 populations. Both populations showed the memory phenotype as shown by a high expression of CD44 and a low expression of CD62L and CCR7 (Fig. 3). However, the α4highβ1+β7+CD4+ T cells expressed lower levels of CCR5 and CXCR3, 2 representative Th1 cell chemokine receptors (1617). The expression of PD-1 and ICOS was also lower in the integrin β1+β7+CD4+ T cells than in β1highCD4+ T cells. Accordingly, integrin α4highβ1+β7+CD4+ T cells secreted smaller amounts of Th1 cytokines such as IFN-γ and TNF-α than integrin α4highβ1highCD4+ T cells (Fig. 4). Furthermore, the integrin α4highβ1+β7+CD4+ T cells secreted small but significant amounts of IL-10. These results suggest that the serosal integrin α4highCD4+ T cells are heterogeneous population with different migratory properties and cytokine secretion.
 | Figure 3Comparison of various activation-related molecules (A) and chemokine receptors (B) among integrin α4highβ1highCD4+ T cells (red line), integrin α4highβ1+β7+CD4+ T cells (blue line), and integrin α4lowCD4+ T cells (green line). PEC and PLC cells obtained from 8-week-old C57BL/6 mice were examined for the expression of indicated cell surface proteins. Data are representative of 12 separate experiments.
|
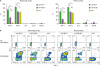 | Figure 4Cytokine production by integrin α4highβ1highCD4+ T cells, integrin α4highβ1+β7+CD4+ T cells, and integrin α4lowCD4+ T cells. (A, B) Peritoneal and pleural cavity cells harvested from 8-week-old C57BL/6 mice were stimulated in vitro with 50 ng/ml PMA and 1.5 mM ionomycin for 4 h. (A) Individual bars represent the percentages of given cytokine-producing cells among α4highβ1highCD4+ T cells (green, α4highβ1high), α4highβ1+β7+CD4+ T cells (purple, α4highβ1+β7+), and α4lowCD4+ T cells (yellowish green, α4low), detected by intracytoplasmic staining of given cytokines. (B) Representative flow cytometric data for given cytokines are shown with or without stimulation. Data are representative of 12 separate experiments.
ns, not significant.
*P<0.05; ***P<0.001.
|
DISCUSSION
Integrin α4β1 (VLA-4) is a principal integrin complex that is essential for T cells to enter the peripheral inflammatory sites such as brain, lung, and pancreatic islets during autoimmune or infectious pathogenetic processes (71819). Integrin α4β1 is not normally expressed on both naïve and memory T cells in the resting condition, implicating a careful regulation of this integrin to prevent excessive infiltration of T cells into peripheral sites. Notably, we previously observed that almost half of peritoneal CD4+ T cells expressed a high level of integrin α4β1 as we designated these cells as integrin α4highCD4+ T cells, which suggests that the PEC is a reservoir of pro-inflammatory T cells (3). In this manuscript, we addressed whether the peritoneal integrin α4highCD4+ T cells expressed other important integrin complexes to gain insights into their functional characteristics and checked whether the PLC also contains this type of T cells. We found that the integrin α4highCD4+ T cells were divided into the major integrin α4highα6+β1highCD4+ T cells and the minor integrin α4highβ1+β7+CD4+ T cells.
The combinations of integrin α and β chains are diverse to form different kinds of adhesion molecules for other cells or extracellular matrix, complement receptor, or receptor for bacterial protein (202122). Integrin α4 and β1 are preferentially expressed on memory T cells rather than naïve T cells. In addition to the pro-inflammatory integrin α4β1, each α4 and β1 integrin chain form alternative combinations that provide additional functional characteristics. Integrin α4 chain is able to combine with integrin β1 or β7, but the probabilities of the combination of α4 chain with 2 β chains are not equal as the integrin β7 is advantageous over the β1 chain in the binding to α4 chain (23). Therefore, the level of the β1 chain expression would determine the expression level of the integrin α4β1 in the presence of a given level of the α4 chain expression. Given the finding that the integrin α4 is highly expressed on the memory phenotype CD4+ T cells, we divided the serosal CD4+ T cells into 3 populations based on the expression of integrin β1 and β7 chains; β1high, β1+β7+, and β1−β7− cells. As β1−β7− CD4+ T cells were integrin α4low cells, integrin α4highCD4+ T cells could be divided into α4highβ1highCD4+ and α4highβ1+β7+CD4+ T cells. Integrin α4highβ1+β7+CD4+ T cells, the smaller population, are thus thought to principally express integrin α4β1 and α4β7. As integrin α4β7 (LPAM-1) is required for the entry into intestine and the pathogenesis of chronic colitis (9), this population appears to have the ability to enter the inflammatory intestinal sites. The role of this peritoneal CD4+ T cell population in gastrointestinal inflammation needs to be addressed in the future.
The β1 integrin is reported to be highly expressed on memory T cells and critical in the maintenance of T cell memory in bone marrow, suggesting that β1 integrin is involved in the entrance of memory T cells into bone marrow (24). Although the integrin α4β1 is likely to be responsible for this migratory behavior, other β1 integrins may be responsible for their unique migration patterns (25). Especially, the very high expression of integrin β1 in the serosal α4highβ1highCD4+ T cells suggested that these cells contained another β1 integrin complex besides α4β1. Integrins α1β1, α5β1, and α6β1 were thought to be good candidates for another β1 integrin expressed on the serosal α4highβ1highCD4+ T cells as these integrins were reported to be expressed on some T cells (142627). Our screening to find the second α integrin partner with the β1 integrin in the α4highβ1highCD4+ T cells resulted in the clear identification of the integrin α6β1 in this population. As the integrin α6β1 is mainly expressed on macrophages and its activity is upregulated by inside-out signaling upon stimulation with PMA (2829), its expression on these innate-like CD4+ T cells is a very interesting feature that may reveal their characteristic migratory pattern such as interstitial migration after transendothelial migration (3031).
In the serosal integrin α4highCD4+ T cells, the expression of integrin α4β7 or α6β1 appeared to be mutually exclusive, arguing that the 2 integrin α4highCD4+ T cell populations are functionally distinctive. As we investigated the activation-related cell surface molecules and cytokine production profiles, we could confirm that the α4highβ1highCD4+ T cells were composed of integrin α4β1+α6β1+α4β7− and α4β1+α6β1−α4β7+ cells. It is interesting to address whether these 2 population originate from the common ancestor population or not. Most importantly, the functional significances of these 2 serosal T cell subsets in diseases such as infection, autoimmunity, cancer, or transplantation need to be investigated in the future. We think that integrin α4 is not a stable marker of these populations as the expression of integrin α4 is altered upon adoptive transfer to Rag-1−/− mice (3) and upregulated by retinoic acid signal (32). The elucidation of developmental markers such as transcription factors is required to follow these cells and human counterpart populations.
In summary, we could identify the distinctive major and minor populations of serosal integrin α4highCD4+ T cells that are different based on the migratory behavior and cytokine secretion. The mutually exclusive expression of integrin α4β7 or α6β1 appear to be an important functional feature of memory or innate T cells determining their preferential pattern of migration.
 XML Download
XML Download