

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
AD
autosomal dominant
CTRL
control
DC
dendritic cell
DOCK8
dedicator of cytokinesis 8
GM-CSF
granulocyte-macrophage colony-stimulating factor
HIES
hyper-IgE syndrome
IFN
interferon
ILC
innate lymphoid cell
ILC1
type 1 innate lymphoid cell
ILC2
type 2 innate lymphoid cell
ILC3
type 3 innate lymphoid cell
PBMC
peripheral blood mononuclear cell
STAT3
signal transducer and activator of transcription 3
TNF
tumor necrosis factor
Treg
regulatory T
INTRODUCTION
Hyper-IgE syndrome (HIES) is a rare primary immunodeficiency disorder characterized by elevated serum IgE, eosinophilia, recurrent skin abscesses, and sinopulmonary tract infections, usually associated with Staphylococcus aureus (1). Most cases of HIES are sporadic, but some familial cases of HIES have been reported, with either an autosomal dominant (AD) or autosomal recessive mode of inheritance. However, the pathogenesis of both the clinical manifestations as well as the immunologic abnormalities remains poorly defined.
Several genetic mutations, such as in signal transducer and activator of transcription 3 (STAT3), tyrosine kinase 2 (Tyk2), and dedicator of cytokinesis 8 (DOCK8), have been implicated in HIES (234). Among these genetic mutations, dominant-negative mutations in STAT3 were identified as a major molecular cause of HIES (2). STAT3 is a transcription factor that binds to the promoter regions of various genes, including those encoding acute-phase proteins. STAT3 is known to play a critical role in signal transduction for many cytokines, including those in the γc family (IL-2, IL-7, IL-9, IL-15, and IL-21), gp130 family (IL-6, IL-11, IL-27, and IL-31), IL-10 family (IL-10 and IL-22), and receptor-type tyrosine kinases (567). Therefore, STAT3 deficiency results in the upregulation of TH1 cytokines, but the downregulation of pro-inflammatory as well as anti-inflammatory responses (2). Moreover, STAT3 has pleiotropic functions in both innate and adaptive immunity (89). A markedly decreased TH17 response is a hallmark of HIES, and STAT3 mutations have been demonstrated to result in the failure of TH17 CD4 cell differentiation (1011). Since IL-17A signaling is involved in neutrophil chemotaxis and proliferation (1213), impaired neutrophil responses due to impaired IL-17A signaling might account for the recurrent staphylococcal infections in patients with HIES (1415).
Moreover, it has been reported that the production of antimicrobial molecules responsible for antifungal responses is tightly regulated by IL-17A (13). In patients with HIES with STAT3 mutations, elevated levels of tumor necrosis factor (TNF)-α, IL-6, and IL-12 in dendritic cells (DCs) were observed (16). Another study showed that tolerogenic DC generation failed in patients with STAT3-deficient HIES due to the disruption of the IL-10 signaling pathway (5). Moreover, increased TNF-α and IL-8 production was detected in HIES peripheral blood mononuclear cells (PBMCs) after stimulation with several concentrations of LPS and/or S. Aureus (17). Finally, hyper-eosinophilia, which is a relatively consistent feature of HIES, has also been observed in mice with myeloid-specific STAT3 deficiency (18). Altogether, these results suggested that patients with STAT3-deficient HIES would have defects in the numbers and function of innate immune cells. Although few studies have been performed to date, the defects in innate immune cells might explain the increased susceptibility to infections in patients with HIES.
Human monocytes can be subdivided based on their expression of the Fcγ receptor III CD16 and lipopolysaccharide co-receptor CD14. “Classical” monocytes account for greater than 70% of total monocytes and are denoted as the CD14+CD16low population (19). Classical monocytes are highly phagocytic, have high myeloperoxidase activity, have high antibody-dependent cell-mediated toxicity, and produce high levels of IL-1, monocyte chemoattractant protein-1 (MCP-1), TNF-α, and IL-6. “Non-classical” monocytes express CD14lowCD16+ antigen and constitute about 10%–15% of blood monocytes. Non-classical monocytes express high levels of major histocompatibility class II receptor, migrate primarily to non-inflamed tissue, and produce high levels of TNF-α, IL-1, IL-6, and IL-12. These cells are more likely to become DCs involved in tissue surveillance. However, the recent identification of an intermediate population identified as CD14+CD16lntermediate supports a new idea that monocytes are a heterogeneous population. Intermediate monocytes are low cytokine producers but express high levels of angiogenic receptors (20). Understanding the different functions of the specific subpopulations of monocytes is necessary for better understanding human diseases.
Innate lymphoid cells (ILCs) have recently been identified as a unique group of innate immune cells (21). ILCs are characterized by a lack of T-cell and B-cell receptors (TCRs and BCRs, respectively), but they rapidly produce a variety of cytokines in response to innate signals, i.e., IL-25, IL-33, thymic stromal lymphopoietin (TSLP), and IL-1α. Traditionally, T-cells were thought as key players in the regulation of the immune response; however, recent emerging studies on ILCs have provided new paradigms for various immune responses, including host defense and tissue homeostasis (212223). To date, the treatment for HIES has relied on prophylactic and therapeutic antibiotics and meticulous skin care, and a general role for ILCs in HIES is still unproven. Because both innate and adaptive responses are important in host defense, we evaluated the frequency and functional response of innate immune cells, including ILCs and monocytes, for the first time in patients with HIES who had recurrent infections.
MATERIALS AND METHODS
Case
PBMCs were taken from a 21-year-old woman who was diagnosed with HIES, and PBMCs from an age-matched healthy control were also used. This study was approved Institutional Review Board of Seoul National University Hospital, which waived the requirement of written informed consent (IRB No. 1707-130-871).
Cell isolation from human blood
PBMCs were diluted with a 3× volume of PBS. Then, 30 ml of diluted blood was carefully layered over 15 ml of Ficoll-Paque in a 50-ml conical tube, followed by centrifugation at 400 ×g for 30 min at 20°C in swinging bucket rotor without a brake. Mononuclear cells at the interface were collected with a transposable pipette. Collected cells were washed with PBS and centrifuged at 300 ×g for 10 min at 4°C. The cell pellets were resuspended in an appropriate volume of buffer and processed for flow cytometry.
Flow cytometry
Single cell suspensions from peripheral blood were stained with the following monoclonal antibodies: PerCP/Cy5.5-anti-CD45 (BD Biosciences, San Jose, CA, USA), fluorescein isothiocyanate (FITC)-anti-CD3 (BioLegend, San Diego, CA, USA), Alexa Flour 700-anti-CD4 (BioLegend, San Diego, CA, USA), Brilliant Violet (BV) 421-anti-CD8(BioLegend, San Diego, CA, USA), FITC-anti-Lineage Cocktail (anti-CD3, anti-CD14, anti-CD19, anti-CD11b, anti-CD11c, anti-CD49b, anti-FcεRIα (BioLegend, San Diego, CA, USA), PE/Cy7-anti-CD127(BioLegend, San Diego, CA, USA), PE-anti-CRTH2(BioLegend, San Diego, CA, USA), BV421-anti-c-kit(BioLegend, San Diego, CA, USA), allophycocyanin (APC)-anti-NKp44(BioLegend, San Diego, CA, USA), PE/Cy7-anti-CD15(BioLegend, San Diego, CA, USA), FITE-anti-CD14(BioLegend, San Diego, CA, USA), and PE-anti-CD16 (BioLegend, San Diego, CA, USA). All samples were blocked with 1 μg Fc block (from 2.4G2 ATCC HB-197) for 15 min before antibody staining at 4°C for 30 min in PBS containing 2% FCS (2% FCS-PBS). Cells were washed twice in 2% FCS-PBS, and then, data were collected on a BD LSRII flow cytometer (BD Biosciences). Analysis of the FACS plots was performed using FlowJo (Treestar, Ashland, OR, USA).
Intracellular cytokine staining
For intracellular cytokine staining, PBMCs were incubated in RPMI (WELGENE, Gyeongsangbuk-do, Korea) containing 10% FBS with PMA (100 ng/ml), ionomycin (1 μg/ml), and BD GolgiStop (BD Biosciences) at 37°C for 4 h. After surface staining, the cells were fixed and permeabilized with BD Cytofix/Cytoperm (BD Biosciences). Finally, the cells were stained with BV421-anti-interferon (IFN)-γ, BV421-anti-granulocyte-macrophage colony-stimulating factor (GM-CSF) (BD Biosciences), APC-anti-IL-5, PE-anti-IL-13, APC-anti-IL-17, and PE-anti-IL-22 (BioLegend). The respective isotype control antibody was also used for each experiment.
RESULTS
Case presentation
A 21-year-old woman was referred to an allergy clinic for the evaluation of multiple skin axillary and buttock abscesses with eosinophilia. She was born full term without perinatal problems. However, she had a history of repeated eczematous rashes on her body and skin abscesses not responding to symptomatic treatment. A diagnosis of HIES was made at 3 months of age based on recurrent skin infections and elevated total serum IgE (773.5 Ku/l) with increased eosinophil counts (38% of white cell blood count, 30,690/μL). She had septic arthritis of the hip joint, abscesses in the liver and lungs, and recurrent multiple cutaneous abscesses caused by methicillin-resistant S. aureus. There was no family history of immunodeficiency. On physical examination at the time of referral, her facial features were coarse with a broad nasal bridge and prominent nose. There was a significant presence of palpable skin lesions on the right axilla and buttock. Bronchial breath sounds were heard in all lung fields, and aeration was decreased in the left lung field. Pulmonary function tests showed severe restrictive patterns. Laboratory tests revealed a WBC count of 4,950/μL, with an elevated eosinophil count (1,100/μl) and total IgE level (158 kU/l). Other laboratory findings, such as liver and renal function tests, urinalysis, and C-reactive protein level, were within normal ranges. A computed tomography scan showed multifocal air cysts or bullae with right necrotic axillary lymph nodes and necrotic abscess in the right lower abdomen, buttock, and groin (Fig. 1). Echocardiography and bone mineral density results were in normal ranges. The clinical and laboratory data were applied to the scoring system of the National Institute of Health, and the total score was 52 points, suggestive of AD-HIES (Table 1). She underwent surgical drainage and excisional biopsy that revealed abscess-forming inflammatory granulation tissue, and was then discharged with prophylactic oral antibiotics (Fig. 2).
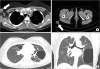 | Figure 1Chest computed tomography scan (A, B). Images showing necrotic lesions in right axillary lymph nodes, subcutaneous tissue of the right buttock (A, white arrows), and multifocal air cysts or bullae of the both lungs (B).
|
Table 1
National Institutes of Health Score for the patient with HIES†

*Asterisks indicate the criteria that the patient satisfied; †Scores of at least 40 points were considered to indicate high likelihood of HIES, whereas scores below 20 indicated low likelihood.
![]()

Impaired type 3 immune responses in a patient with HIES
Several studies have measured the population of T cells in the PBMCs of patients with HIES; however, the results varied according to the experimental conditions. Therefore, we first evaluated the T cell populations in the PBMCs of a patient with HIES (n=1) and age-matched controls (n=3) by flow cytometry. The percentages of CD3+ T cells in the PBMCs were similar between the patient with HIES and the controls (Fig. 3A and B). We further analyzed the T cell subpopulations and found that the percentages of CD4 and CD8 T cells were within normal ranges, but approximately 1.9 times more Foxp3+ regulatory T (Treg) cells were detected in the PBMCs of the patient with HIES than in those from the controls (Fig. 3A and B). Similar to our results, Heimall et al. (1) reported that normal numbers of CD4 and CD8 T cells existed in the PBMCs of patients with HIES. Additionaly, a marked decrease in the TH17 response has been reported as a hallmark of HIES due to STAT3 mutations (24). Therefore, we stimulated PBMCs with PMA/ionomycin and then measured the production of various cytokines, including IL-17A from T cells. Type 2 cytokine expression in T cells was similar between the patient with HIES and the control subjects. However, expression of type 1 and type 3 cytokines such as IFN-γ, IL-17 and IL-22, respectively, in T cells was lower in the patient with HIES than in the controls (Fig. 3C and D). These findings suggested that our patient with HIES may have a defective STAT3 signaling pathway, which resulted in reduced type 3 cytokine production from T cells.
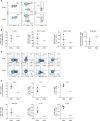 | Figure 3The distribution of T cells in PBMCs from healthy controls and a patient with HIES. (A, B) CD4+ T cells, CD8+ T cells, and Treg cells were analyzed by flow cytometry. (A) Representative flow cytometric profiles of the T cell population. (B) Graphs show the mean percentages of CD3+ T cells, CD4+ T cells, CD8+ T cells, and Treg cells±SEM (n=3). (C, D) Intracellular IFN-γ, IL-5, IL-13, GM-CSF, IL-17, and IL-22 expression in T cells was analyzed by flow cytometry. (C) Representative flow cytometric profiles of cytokine expression by T cells. (D) Graphs show the mean percentage of positive cells±SEM (n=3).
CTRL, control; GM-CSF, granulocyte-macrophage colony-stimulating factor; HIES, hyper-IgE syndrome; IFN, interferon; PBMC, peripheral blood mononuclear cell; SEM, standard error of the mean; Treg, regulatory T.
|
Altered function of ILCs in a patient with HIES
There have been several reports regarding the roles of adaptive immune responses in the pathogenesis of HIES; however, the roles of innate immune responses in HIES remain unknown. For example, Gutierrez-Hincapié et al. (25) measured the number and frequency of innate immune cells in PBMCs from patients with HIES, but there were no differences in terms of the absolute number of leukocytes, neutrophils, monocytes, mDCs, pDCs, and NK cells in PBMCs from patients with HIES. However, they found that the number of invariant NK T (iNKT) cells was significantly reduced in the AD-HIES group; hence, these results indicated that innate immune responses were also impaired in the patients with HIES.
As an innate counterpart of TH cells, ILCs play important roles by producing types 1, 2, and 3 cytokines. However, the role of ILCs in HIES has not been reported. To determine the role of ILCs in our patient with HIES, we analyzed ILCs using flow cytometry. Compared to the control subjects, the percentage of total ILCs (Lineage−CD127+) was lower in the PBMCs from the patient with HIES (Fig. 4A). Next, we further analyzed the subset of ILCs by adding several surface markers such as CRTH2, c-kit, and NKp44 to distinguish type 2 innate lymphoid cells (ILC2s; Lineage−CD127+CRTH2+), type 3 innate lymphoid cells (ILC3s; Lineage−CD127+CRTH2−c-kit+), and type 1 innate lymphoid cells (ILC1s; Lineage−CD127+CRTH2−c-kit−). Interestingly, most of ILCs in the patient with HIES were ILC1s and not ILC2s or ILC3s (Fig. 4A and B). Intracellular staining of ILCs also revealed that IL-17A and IL-22 (type 3) cytokine production in the PBMCs was lower in the patient with HIES than in the controls, but IFN-γ production was similar (Fig. 4C and D). These results indicated that ILCs, as well as T cells, had altered functions in the patient with HIES.
 | Figure 4The distribution of ILCs in PBMCs from healthy controls and a patient with HIES. (A, B) ILC1s (CD45+Lin−CD127+CRTH2−c-kit−NKp44−), ILC2s (CD45+Lin−CD127+CRTH2+), and ILC3s (CD45+Lin−CD127+CRTH2−c-kit+) were analyzed by flow cytometry. (A) Representative flow cytometric profiles of ILC subsets. (B) Graphs show the mean percentage of total ILCs, ILC1s, ILC2s, and ILC3s±SEM (n=3). (C, D) Intracellular IFN-γ, IL-5, IL-13, GM-CSF, IL-17, and IL-22 expression in ILCs was analyzed by flow cytometry. (C) Representative flow cytometric profiles of cytokine expression in ILCs. (D) Graphs show the mean percentage of positive cells±SEM (n=3).
CTRL, control; GM-CSF, granulocyte-macrophage colony-stimulating factor; HIES, hyper-IgE syndrome; ILC, innate lymphoid cell; PBMC, peripheral blood mononuclear cell; SEM, standard error of the mean.
|
CD14+CD16intermediate monocytes are increased in a patient with HIES
To test the proportion of monocytes in the patient with HIES, we analyzed monocyte subsets using flow cytometry and antibodies to CD14 and CD16. The dot plot of the data indicates that was a large increase in monocytes with CD14 expression in the PBMCs of the patient with HIES compared with the normal controls (Fig. 5A and B). Interestingly, CD14+CD16lntermediate monocytes were higher in the patient with HIES than in the controls, while there was no significant difference in the proportion of classical monocytes (CD14+CD16low) and non-classical monocytes (CD14lowCD16+).
 | Figure 5The distribution of monocytes in PBMCs from healthy controls and a patient with HIES. (A, B) Monocytes (CD45+CD15−CD14+) were analyzed by flow cytometry. (A) Representative flow cytometric profiles of monocyte populations. (B) Graphs show the mean percentages of monocytes, classical monocytes (CD14+CD16low), intermediate monocytes (CD14+CD16intermediate), and non-classical monocytes (CD14lowCD16+)±SEM (n=3). (C) The correlations between ILC1s and classical monocytes, intermediate monocytes, and non-classical monocytes were analyzed in healthy controls (black) and a patient with HIES (red).
CTRL, control; HIES, hyper-IgE syndrome; ILC, innate lymphoid cell; SEM, standard error of the mean.
|
Recently, Xiong et al. (26) reported on the crosstalk between inflammatory monocytes and ILCs. The authors suggested that IL-17A from ILCs could enhance the antimicrobial activities of inflammatory monocytes. To evaluate the correlation between monocytes and ILCs, we tested the association between subsets of monocytes and ILCs from healthy control and a patient with HIES (Fig. 5C). Although the number of samples was limited, the proportion of CD14+CD16lntermediate monocytes had a positive trend with an increase in ILC1s. In contrast to CD14+CD16lntermediate monocytes, non-classical monocytes (CD14lowCD16+) had a positive trend with an increase in ILC2s and ILC3s, but not ILC1s (Fig. 5C, Supplementary Fig. 1).
Taken together, these results indicated that innate immune cells are indeed altered in this patient with HIES; therefore, further studies to define the functional changes in innate immune cells during disease progression are needed.
DISCUSSION
HIES is a complex primary immunodeficiency disorder characterized by recurrent staphylococcal skin abscesses and pneumonia, atopic dermatitis, and extremely high serum IgE levels. Recent studies have shown that dominant-negative mutations of the STAT3 gene are a major molecular cause of HIES in most patients (227). However, our understanding of the immune alterations underlying HIES remains limited. In this report, we focused on the innate and adaptive immunological abnormalities in HIES.
The STAT3 signaling pathway was originally discovered in the context of components downstream of the IFN-α, IFN-γ, and IL-6 signaling pathways (2829). STAT3 is not only crucial for transducing signals, but it also functions as a critical transcription factor that regulates the expression of a wide range of genes. Therefore, STAT3 plays a crucial role in the function of various cell types, including stromal cells, tumor cells, epithelial cells, and immune cells. The susceptibility to infection observed in patients with HIES can be explained by the lack of IL-17A production from T cells due to impaired STAT3 signaling. In addition, impaired production of IFN-γ and TNF-α by T cells (30), diminished memory T-cell populations, and decreased delayed-type-hypersensitivity responses have variably been described in patients with HIES (15).
Although the functional alteration of adaptive immunity, such as changes in T cells and B cells, has been relatively well studied, innate immunological abnormalities that can explain the unique susceptibility to particular infections seen in HIES have not been identified. In this regard, we evaluated the innate immune cells observed in a HIES case. Of interest, the percentage of total ILCs in PBMCs was lower in the patient with HIES than in the controls, while the percentage of T cells remained relatively similar. Moreover, our data also indicated that ILC3s in PBMCs were significantly lower in the patient with HIES than in the controls. Although STAT3 is known for IL-17A production in adaptive arms, Guo et al. (31) reported that STAT3 expression in RORγt+ ILC3s, but not in T cells, was essential for protection against Citrobacter rodentium. They also suggested that transcriptional regulation of ILC3s, TH17, and TH22 cells might differ. Therefore, the functions of TH17 and ILC3s in HIES would be different, although they all produce type 3 cytokines and are regulated by STAT3. Thus, the mechanisms underlying the differential regulation by STAT3 in ILCs and T cells need to be further investigated.
In a current study, we also observed an increased number of intermediate monocytes in the PBMCs of a patient with HIES. In clinical settings, intermediate monocytes (CD14+CD16lntermediate) have been noted to increase in several diseases (32). For example, in patients with moderate and severe forms of asthma, a pronounced increase in intermediate monocytes was observed (19). In patients with rheumatoid arthritis, a strong increase in intermediate monocytes but a decrease in classical monocytes was observed (33). Finally, in patients with colorectal cancer, the percentage of intermediate monocytes was found to be increased, and this phenomenon was more pronounced in local than in metastatic disease (34). Altogether, these results show that intermediate monocytes are increased in several diseases; however, the roles of intermediate monocytes in the pathogenesis of disease still remain unclear. Moreover, there has been no report on the roles of monocytes in HIES except that IL-10 signal transduction was found to be defective in the monocyte derived DCs of patients with HIES (5). In terms of the effect of IL-10 on the function of monocyte, Skrzeczyńska-Moncznik et al. (35) reported that the highest IL-10 production was detected in intermediate monocytes in response to LPS and to zymosan. Extending this concept, Tsukamoto et al. (36) found that classical monocytes (CD14brightCD16−) became intermediate/non-classical monocytes (CD14brightCD16+) when they were stimulated with IL-10. Interestingly, we also observed that intermediate monocytes were increased in the PBMCs of a patient with HIES, and the increase in intermediate monocytes might have a suppressive effect of excessive immune responses. In our patient with HIES, we observed that intermediate monocytes negatively correlated with ILC3s. If we can demonstrate that intermediate monocytes interact with a specific type of ILC and regulate inflammation in several disease conditions, it would support these novel and biologically meaningful concepts. However, further studies are required to fully elucidate this idea.
Taken together, our results suggest several important findings regarding HIES. First, our patient with HIES had few defects in peripheral T cells, but certain changes were observed in the innate immune cells, including ILCs and monocytes. Second, our data showed that type 3 and type 1 cytokine production from ILCs, which are essential for the response to acute bacterial infections, were significantly reduced in the PBMCs of the patient with HIES. Finally, the percentage of monocytes, especially intermediate monocytes, was significantly increased the PBMCs of the patient with HIES. Recent studies have suggested that intermediate monocytes play a regulatory role by producing IL-10. Therefore, a better understanding of the molecular mechanisms of intermediate monocytes is required.
Currently, the treatments for patients with HIES are primarily focused on controlling recurrent infections, such as bacterial and fungal infections. Moreover, previous studies have suggested the benefits of bone marrow transplantation, Ig replacement, and administration of IFN-γ and granulocyte colony-stimulating factor (G-CSF) for HIES treatment (37). However, our results indicate the possibility that the “innate arms” also play critical roles in patients with HIES. To improve the long-term quality of life of patients with HIES, it is necessary to develop a new treatment strategy based on both innate and adaptive immune mechanisms.
 XML Download
XML Download