

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Acute-phase serum amyloid A (SAA) is primarily expressed in the liver in response to inflammation or infections (12). Recently, extrahepatic expression of SAA has been elucidated in many tumor (3), inflammatory (4), obese (56), and normal tissues (2). Human and mouse have 4 SAA isoforms (SAA1, 2, 3, and 4) (7). In human, SAA1 and SAA2 are mainly synthesized during acute-phase inflammatory reactions (8) and share approximately 95% of overall sequence identity in their promoter regions, exons, and introns (9). On the other hand, human SAA3 is pseudogene and SAA4 is constitutively expressed. Human SAA1/2 is comparable to mouse SAA3 which is expressed abundantly and extrahepatically (10). SAA is released in response to proinflammatory cytokines such as IL-6, IL-1, and tumor necrosis factor-α (TNF-α). The concentration levels of SAA during inflammation reach up to 1,000-fold greater than those in the non-inflammatory state.
Reproductive processes such as ovulation, menstruation, implantation, and labor are comparable to inflammatory processes (111213). Inflammation is an active process which involves the release of inflammatory cytokines, chemokines, and peptide growth factors. All of reproductive events involve upregulation of inflammatory mediators such as cytokines, and growth factors (11). In particular, cytokines expressed locally in the granulosa cells and endometrial tissue cause ovulation and implantation, respectively. Moreover, ovarian hormones are likely to be mediators of inflammatory process (11). Furthermore, nonsteroidal anti-inflammatory agents such as indomethacin inhibit ovulation in mammalian species (14).
Our previous study revealed that TNF-α specifically and abundantly increased SAA3 among acute-phase proteins in mouse granulosa cells (15). TNF-α, a multi-functional hormone-like polypeptide, is well known to modulate many genes involved in inflammation, infection, and malignancy. In particular, ovarian TNF-α locally regulates steroidogenesis, folliculogenesis, ovulation, and fertility (16171819). Although SAA3 is the most abundant gene induced by TNF-α in granulosa cells (715), the ovarian localization of SAA3 is unknown. Thus, the present study was designed to investigate the localization of SAA3 in the mouse ovary using both a mouse ovulation model and model of inflammatory condition.
MATERIALS AND METHODS
Animal handling
C57BL6 female mice from Harlan, Inc. (Indianapolis, IN, USA) were used from breeding colonies in our laboratory. All handling of animals and procedures were approved by the Institutional Animal Care and Use Committee at the University of Kansas Medical Center.
Mouse ovulation model and ovarian inflammatory condition
For mouse ovulation model, mice were subcutaneously (sc) injected with 2.5 IU equine chorionic gonadotropin (eCG; Sigma-Adrich, St. Louis, MO, USA) on day 25 to initiate follicle development for 48 h. Fifty hours after eCG, 2.5 IU human chorionic gonadotropin (hCG; Sigma-Adrich) was administered sc to induce ovulation and luteinization by 72 h (Fig. 1). Mice were euthanized at the indicated time beginning on day 25 through day 28 and ovaries were collected for 10 μm frozen sectioning and in situ hybridization. For ovarian inflammatory condition, 2 months old mice were treated intraperitoneally with IL-1β (1 μg/mouse), were euthanized at 2 h post-injection and ovaries were collected for frozen sectioning and in situ hybridization.
 | Figure 1Schematic of mouse ovulation model and ovarian localization of CYP19 mRNA. Ovaries were collected at 0, 24, 48 h after eCG (2.5 IU) to initiate follicular development, and 2, 24 h after hCG (2.5 IU) to induce ovulation and luteinization. The mouse ovulation model is characterized by the expected time- and cell-specific expression of CYP19 mRNA using in situ hybridization with digoxigenin CYP19 RNA probe.
|
Digoxigenin CYP19 and SAA3 RNA probes
Total RNA was isolated using TRIzol® reagent (Invitrogen, Grand Island, NY, USA). The reverse transcription (RT) reaction conditions, using random primers with M-MLV (Invitrogen), were at 42°C for 60 min, followed by 94°C for 10 min. Specific primers for mouse CYP19 aromatase and SAA3 were designed as follows: 5′-TCA ATA CCA GGT CCT GGC TA-3′ (forward) and 5′-TGC TTG ATG GAC TCC ACA CA-3′ (reversed) for CYP19; and 5′-AGC CTT CCA TTG CCA TCA TTC TT-3′ (forward) and 5′-AGT ATC TTT TAG GCA GGC CAG CA-3′ (reversed) for SAA3. PCR was performed in 30-cycle increments under the following conditions: denaturation at 94°C for 1 min; annealing at 58°C for 1 min; and extension at 74°C for 1 min. Amplified PCR product (374 bp) was analyzed by electrophoresis in 2% agarose gels containing 1 µg ethidium bromide/ml and was purified by a spin-column (Gel Extraction System; Qiagen, Valencia, CA, USA) to absorb the DNA. The PCR products were inserted into the pGEM®-T vector (Promega, Madison, WI, USA). Antisense and sense linear template DNAs were made by digestion with NcoI and SpeI enzymes, respectively, according to inserted orientation. Linear template DNAs were labeled with digoxigenin-UTP (Roche, Indianapolis, IN, USA) by in vitro transcription with SP6 (antisense) and T7 (sense) RNA polymerase as described previously (20).
In situ hybridization
Nonradioactive methods for in situ hybridization on frozen sections were performed using digoxigenin-labeled CYP19 and SAA3 RNA probes (antisense and sense) as described previously (20).
Data analysis from Gene Expression Omnibus (GEO) dataset
Data analysis was performed using microarray data sets deposited in the National Center for Biotechnology Information (NCBI) GEO (https://www.ncbi.nlm.nih.gov/geo/) database under accession number GSE23084 and GDS1677. We employed Gitools 2.2.3 (Biomedical Genomics Group, Barcelona, Spain; http://www.gitools.org) based on Oracle Java 7, an open-source tool to perform Genomic Data Analysis and Visualization as interactive heat-maps (22) as described previously (23).
RESULTS AND DISCUSSION
The localization of SAA3 mRNA in the whole ovary was performed in mouse ovulation model using in situ hybridization as described previously (720). Ovaries were collected at post-eCG injection (0, 24, and 48 h) and post-hCG (2 and 24 h) (Fig. 1). First, ovarian CYP19 (aromatase) mRNA was localized to characterize and validate the mouse ovulation model (Fig. 1). CYP19 mRNAs are expressed in the granulosa layer of large follicles during follicular development, with maximal expression occurring immediately after ovulation and loss of expression with luteinization (Fig. 1) as anticipated (2425). The expression pattern of ovarian CYP19 mRNA indicates that the mouse ovulation model reflects the physiological process of follicular development and ovulation. Next, we determined the localization of SAA3 mRNA expression in the whole ovaries using the mouse ovulation model. We confirmed no signal with sense SAA3 probe (Fig. 2). SAA3 mRNA is lightly and broadly expressed throughout the whole ovary even prior to eCG injection (Fig. 2). During follicle development, granulosa cells express higher SAA3 mRNA than thecal-interstitial cells, and small follicles express higher SAA3 mRNA than large follicles (Fig. 2). Interestingly, SAA3 mRNA is intensely expressed in ovarian follicle atresia (Fig. 2) in which immature ovarian follicles are degenerated by granulosa cell apoptosis. Because atresia in ovarian follicles involves atretogenic factors such as TNF-α and Fas ligands (26), these factors are likely to induce SAA3 mRNA which is strictly regulated by NF-κB signaling (715). At 72 h ovary (48 h post-eCG and 24 h post-hCG injections), corpora lutea (CL) contain cells with intense SAA3 mRNA hydridization (Fig. 2). Because TNF-α is a product of luteal macrophages that infiltrate into the CL (27), immune cell-derived proinflammatory cytokines are likely to induce SAA3 during CL formation and luteal regression. NF-κB mediated signaling is primary pathway inducing SAA3 expression in mouse granulosa cells (715). Therefore, we employed treatment with IL-1β a proinflammatory cytokine activating NF-κB signaling, and assessed localization of SAA3 mRNA by in situ hybridization. Intense SAA3 mRNA hybridization was localized to CL (Fig. 3) similar to the pattern observed in newly formed CL using the ovulation model (Fig. 2). IL-1β-treated ovary has more intense and extensive SAA3 mRNA expression compared to non-treated ovary (Fig. 3). Interestingly, following IL-1b treatment SAA3 mRNA was intensely localized to the thecal-interstitial cell layers in both small and large follicles (Fig. 3). These findings indicate that ovarian follicles likely protect their oocytes by blocking harmful effects due to inflammatory stimulation. Direct effects of proinflammatory cytokines on SAA3 expression were then assessed using cultured primary granulosa cells. In vitro treatment with TNF-α, IL-1α, or IL-1β strongly induces SAA3 mRNA expression in mouse granulosa cells compared to non-treated granulosa cells (Fig. 4A). Granulosa cell-specific expression of SAA3 was further assessed by collection of granulosa throughout the induction of follicle development and ovulation in the mouse ovulation model. Granulosa cells collected at 48 h post-eCG plus 24 h post-hCG injection highly express SAA3 mRNA (Fig. 4B), reflecting CL derived SAA3 expression as indicated in Fig. 2. On the other hand, granulosa cells collected prior to this time point expressed modest SAA3 documented by only a faint band following RT-PCR. SAA3 expression in immature granulosa cells could be stimulated by LPS, a strong inflammatory mediator. In another set of experiments mice were injected intraperitoneally with LPS and then treated with gonadotropins as the ovulation model with granulosa cells collected at various time points. Granulosa cells collected following injection with LPS and prior to eCG injection express increase SAA3 mRNA compared to mice not receiving LPS (Fig. 4B). In addition, SAA3 mRNA expression was further increased in cells collected at the 48 h post-eCG plus 24 h post-hCG time point in LPS treated mice compared to mice not receiving LPS (Fig. 4B). SAA expression in mouse ovaries was further assessed by examination of 2 different GEO data sets. Each data set indicates that SAA3 is the primary SAA isoform in the ovary (Fig. 4C and 4D). Furthermore, SAA3 expression was increased following eCG plus 24 h post hCG (Fig. 4C) similar to the findings of the current study (Fig. 4B). Additional data indicate that SAA3 expression in granulosa cells comprising the cumulus-oocyte-complex is increased following eCG plus 16 h post hCG (Fig. 4D). Together these 2 GEO datasets support the findings of the current study on high expression of SAA3 in CL as shown in Fig. 2.
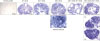 | Figure 2Localization of SAA3 mRNA in the mouse ovary during gonadotropin-induced follicle development and ovulation. Hybridization of antisense SAA3-labeled riboprobes was visualized as purple-blue precipitate in the whole ovary at respective time points (0, 24, 48 h post-eCG injection, and 2, 24 h post-hCG injection). Incubation of tissue sections with labeled sense probe revealed no hybridization.
|
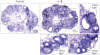 | Figure 3IL-1β treatment in vivo induced SAA3 mRNA in the mouse ovary. Adult cycle mice (2 months old) were treated intraperitoneally with IL-1β (1 μg/mouse). Mice were euthanized at 2 h post-injection and ovaries were collected for 10 μm frozen sectioning and in situ hybridization.
|
 | Figure 4Expression of SAA3 mRNA in mouse granulosa cells. (A) Effect of proinflammatory cytokines on SAA3 mRNA in mouse granulosa cells. Mouse granulosa cells were incubated with vehicle, TNF-α (10 ng/mL), IL-1α (10 ng/mL), and IL-1β (10 ng/mL) for 2 h. RT-PCR was performed using SAA3 primers. L19 was used as a loading control. (B) Expression of SAA3 mRNA in mouse granulosa cells collected from ovaries of mice during the ovulation model and after 2 h post-LPS (100 μg/mouse) challenge. (C) Expression levels of SAA isoforms in mouse granulosa cells based on the NCBI GEO dataset (GSE23084). (D) Expression levels of SAA isoforms in cumulus oocyte complex based on the NCBI GEO dataset (GDS1677).
NCBI, National Center for Biotechnology Information; M, molecular marker in base pairs; Con, control.
|
In conclusion, SAA3 mRNA is broadly expressed by in situ hybridization throughout the whole ovary. In particular, atretic follicles and CL express intensely elevated levels of SAA3 mRNA. In addition, proinflammatory conditions such as treatment with LPS or IL-1β potentiate SAA3 mRNA expression in CL and in thecal-interstitial layers around follicles. Further study will be required to identify the specific cell type expressing SAA3, however, these results indicate that SAA3 is likely to play a role in ovarian follicular atresia and luteal formation and regression.
 XML Download
XML Download