

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
C5a
complement 5a
C5aR
complement 5a receptor
C5aRA
C5aR antagonist
C5aR KO
C5aR knock-out
cAMP-PKA
cyclic adenosine monophosphate-dependent protein kinase
CIN
cefsulodin-irgasan-novobiocin
E
enterocyte
GP2
glycoprotein 2
LPS
lipopolysaccharide
M
microfold
MLN
mesenteric lymph node
p-C5aR
phosphorylated C5aR
PKA
protein kinase A
PP
Peyer's patch
p-PKA
phosphorylated-PKA
TLR
Toll-like receptor
WT
wild-type
INTRODUCTION
The gut mucosal epithelial cells play a regulatory role as a barrier function to enormous amount of microbiota (12). In addition, influx of luminal Ags is also tightly regulated by specialized mucosal epithelial cells including microfold cells (M cells), goblet cells, and CX3CR1+ lamina propria cells (3). Among them, M cells located in follicle-associated epithelium (FAE) of Peyer's patches (PPs) are specialized epithelial cells for uptake of luminal Ags (4). M cells have distinguished structural features such as short and irregular microvilli and a high capacity of transcytosis which enables the influx of various sizes of substance ranged from microparticles to bacteria (5). Although M cells play a role as gateway for Ag sampling, they are, at the same time, exploited as a primary target for the infection of various pathogens such as Salmonella typhimurium, Shigella flexneri, Yersinia entericolitica, and Brucella abortus for across the intestinal epithelium (6). Given that apical area of M cells has several receptor proteins such as glycoprotein 2 (GP2) which can interact with FimH+ bacteria, receptor-mediated infection is one of the plausible mechanisms of pathogen exploit the M cells (7). Additionally, complement 5a (C5a) receptor (C5aR or CD88) is stimulated by Y. enterocolitica and B. abortus utilizes cellular prion protein as an invasive receptor (78). However, it is still unclear how the pathogens manipulate M cells for their infection.
Several recent studies suggest that host cell receptor crosstalk can be manipulated by microbes (9). For example, Staphylococcus aureus blocks inflammatory response mediated by Toll-like receptor (TLR) 2 signaling through activation of immunoreceptor tyrosine-based inhibitory motif (ITIM)-coupled receptors (10). Additionally, some microorganisms including Helicobacter pylori, Mycobacterium spp., and Candida albicans bind to both dendritic cell-specific ICAM3-grabbing non-integrin (DC-SIGN) and TLRs and then induce the IL-10 expression which, in turn, inhibits the TLR-mediated signaling to induce inflammation (111213). C5aR is also closely associated with microbe-mediated manipulation of the signal crosstalk (9). For instance, Porphyromonas gingivalis interacts with TLR1/2 and TLR4 and, at the same time, C5a released by the bacterial Arg-specific cysteine proteinase-mediated cleavage enhances the intracellular Ca2+ signaling via C5aR activation (14). This crosstalk between C5aR and TLRs inhibits the inducible nitric oxide synthase (iNOS)-dependent killing of the bacteria. Given that TLRs and C5aR exist in M cells, this manipulation of signal crosstalk may also occur in M cells, although it is not studied yet (81516).
In this study, we tried to characterize the possible manipulation mechanism of signal crosstalk between C5aR and TLRs using M cell-exploiting pathogen, Y. enterocolitica. Initially, we found the TLRs co-localize with C5aR on apical area of M cells. In addition, it was identified that Y. enterocolitica infection into M cells regulated crosstalk between C5aR and TLRs via cyclic adenosine monophosphate-dependent protein kinase A (cAMP-PKA) signaling. Finally, we confirmed that the infection of Y. enterocolitica into M cells is critically regulated by C5aR both in microfold (M)-like cells and C5aR knock-out (C5aR KO) mice. Collectively, we suggest that exploit the crosstalk between C5aR and TLR signaling is one of infection mechanisms utilized by mucosal pathogens to infect M cells.
MATERIALS AND METHODS
Mice and experimental materials
All chemical and experimental materials were purchased from Sigma Chemical Co. (St. Louis, MO, USA), unless otherwise specified. Syngeneic BALB/c and C5ar1tm1Cge KO mice were purchased from Charles River Technology through Orient Bio (Sungnam, Korea). NKM 16-2-4 Ab and anti-GP2 Ab were purchased from MBL Life Science (Nagoya, Japan). Anti-C5aR Ab, anti-phosphorylated C5aR (p-C5aR) Ab, anti-TLR1/2 Ab, and anti-TLR4 Ab were purchased from Abcam (Cambridge, MA, USA). Anti-phosphorylated-protein kinase A (p-PKA) substrate Ab was purchase from Cell Signaling (Boston, MA, USA). Yersinia enterocolitica (ATCC 23715) used in this research was obtained from Korean Culture Center of Microorganisms (KCCM, Seoul, Korea).
Immunofluorescence analysis
Whole mouse PPs were fixed with 4% paraformaldehyde for 2 days, stained with each indicated Ab followed by the appropriate fluorescent dye-conjugated secondary Abs. The samples were analyzed by confocal laser scanning microscopy (LSM 510 META; Carl Zeiss, Thornwood, NY, USA).
To monitor the localization of Y. enterocolitica, bacteria were washed in PBS and then incubated for 10 min at room temperature in PBS containing 5 μM CFSE Kit (Molecular Probes, Eugene, OR, USA). After washing with PBS, concentration of bacteria was determined by spectrophotometer (Biorad, Hercules, CA, USA). The whole mount PP prepared from mice orally administered with CFSE-labeled Y. enterocolitica (1×108 bacteria) were stained with anti-p-C5aR, anti-TLR1/2, and Alexa Fluor 350-conjugated wheat-germ agglutinin (WGA; Molecular Probes). The whole-mount samples were analysed by confocal laser scanning microscopy.
To analyse the PKA activation, PP samples were prepared from the mice orally administered with CFSE-labeled Y. enterocolitica (1×108 bacteria), fixed, frozen sectioned, and stained with anti-GP2 Ab and anti-p-PKA substrate Ab and then counter stained with DAPI (Molecular Probes). The slide samples were analysed with confocal laser scanning microscopy.
Quantitative real-time PCR
A human colon carcinoma cell line, Caco-2, and a human Burkitt's lymphoma cell line, Raji, were purchased from the American Type Culture Collection (Manassas, VA, USA). The in vitro human M-like cells which had been prepared as described in our previous study were treated with recombinant human C5a (R&D Systems, Minneapolis, MN, USA), lipopolysaccharide (LPS) and C5a, or Y. enterocolitica for 6 h and then total mRNA was purified using the RNeasy Plus mini kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol (17). The transcript levels of each indicated genes were monitored by human cAMP/Ca2+ signaling pathway array kit (PAHS-066Z; Qiagen) according to the manufacturer's instructions using an Applied Biosystems 7500 Real-Time PCR Instrument System (Life Technology, Carlsbad, CA, USA).
Intracellular survival assay
The viability of internalized Y. enterocolitica was monitored by an antibiotic protection-based intercellular survival assay (14). Briefly, inverted cultured M- and enterocyte (E)-like cells were allowed to be infected by Y. enterocolitica (multiplicity of infection [MOI]=1:1) with or without C5aR antagonist (C5aRA) for 2 h at 37°C. The cells were followed by washing to remove extracellular non-adherent bacteria and 2 h treatment with antibiotics (20 µg/mL tetracycline, 25 µg/mL kanamycin, and 20 µg/mL chloramphenicol) to eliminate residual or extracellular adherent bacteria. The M- and E-like cells were subsequently cultured for 2 h. Finally, the cells were washed and lysed in sterile distilled water and viable counts of internalized Y. enterocolitica were determined by plating serial dilutions of cell lysates on cefsulodin-irgasan-novobiocin (CIN) agar plates (Beckton Dickinson, Franklin Lakes, NJ, USA) which were incubated in 37°C chamber.
In vivo infection of Y. enterocolitica
After oral infection of 1×108 colony-forming unit (CFU) of Y. enterocolitica into wild-type (WT) and C5aR KO mice, the number of survived Y. enterocolitica in PPs and mesenteric lymph nodes (MLNs) were measured by plating serial dilutions of tissue homogenates on CIN agar plates which were incubated in 37°C chamber.
Statistical analysis
Statistical results were analyzed using Prism 7 (GraphPad, La Jolla, CA, USA). Data are presented as means±standard errors (SEs) of 3 individual mice per group. Differences in the means of multiple independent variance were determined by one-way ANOVA, followed by Tukey's post hoc test.
RESULTS
TLRs are co-localized with C5aR on the apical area of M cells
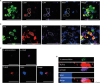
Our previous study reported that C5aR interacts with Y. enterocolitica on apical area of M cells but we did not analyze its implication on the process of Y. enterocolitica infection (8). In order to understand if Y. enterocolitica exploit C5aR for their infection via crosstalk between C5aR and TLRs as a novel immune subversion mechanism, we first identified the expression of TLR1/2 and TLR4 on M cells expressing C5aR (Fig. 1A and 1B). Although the expression level of TLR1/2 and TLR4 was not high, they co-localized with C5aR on M cells in whole-mounted PPs. In order to further confirm the expression of TLR1/2 during the Y. enterocolitica infection, PPs were prepared from WT mice which had been orally infected with CFSE-labeled Y. enterocolitica. Importantly, it was found that TLR1/2 expression was detected together with p-C5aR and CFSE-labeled Y. enterocolitica on PP M cells (Fig. 1C and 1D). Collectively, it was conceivable that Y. enterocolitica infection into M cells is associated with both C5aR and TLRs and may exploit signal crosstalk between C5aR and TLRs.
Figure 1
Y. enterocolitica interacted with C5aR and TLR1/2 in PP M cells. (A, B) Whole-mounted PPs were stained with either NKM 16-2-4 anti-M cell Ab (A) or anti-GP2 Ab (B), anti-C5aR Ab, either anti-TLR4 Ab (A) or anti-TLR1/2 Ab (B), and WGA. (C, D) PPs were prepared at 10 min after oral administration of CFSE-labeled Y. enterocolitica and stained with anti-p-C5aR Ab, anti-TLR1/2 Ab, and WGA. The samples were analyzed by confocal laser scanning microscopy. (C) and (D) show stacked images of XY and XZ images. The dotted lines depict M cells. Scale bars represent 20 μm.
WGA, wheat-germ agglutinin.
Y. enterocolitica induces the cAMP-PKA signaling in M cells
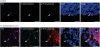
We investigated whether Y. enterocolitica infection induces the enhancement of cAMP-associated gene expression in M-like cells because signal crosstalk between C5aR and TLRs increase the intracellular cAMP level (14). The human M-like cells were prepared from Caco-2 cells by co-culturing with Raji cells and then they were stimulated with either C5a, C5a together with LPS, or Y. enterocolitica. When we monitored the change of the gene expression through quantitative real-time PCR, the expression level of certain genes including Ddit3, Eno2, Pou2af1, and S100a12 was enhanced significantly (p<0.01 for Eno2 and p<0.001 for Pou2af1 and S100a12) by treating the M-like cells either with C5aR together with LPS or with Y. enterocolitica compared to that only with C5a treatment (Fig. 2). Given that those genes are regulated by calcium responsive elements or calcium responsive element enhancer, it suggests that treatment of M-like cells either with C5a together with LPS or Y. enterocolitica infection into M-like cells increases the intracellular calcium level. To further confirm this speculation, cAMP-PKA, PKA, activation was monitored in PP M cells of the mice orally administered with CFSE-labeled Y. enterocolitica because the increased intracellular calcium level activates PKA signaling through the increase of cAMP level (Fig. 3) (18). It was shown that PKA substrate was phosphorylated in GP2+ M cells prepared from the mice orally administered with Y. enterocolitica. Importantly, the similar phosphorylation of PKA substrate was not detected in GP2+ M cells prepared from control mice without oral administration of Y. enterocolitica. Consequently, we confirmed the crosstalk between C5aR and TLRs in M cells by Y. enterocolitica infection and the infection regulates the cAMP-PKA signaling in M cells.
Figure 2
The expression level of cAMP signaling-related genes increased in human M-like cells incubated with Y. enterocolitica. The human M-like cells were differentiated from Caco-2 cells by co-culturing with Raji. After stimulating the cells with either C5a together with LPS, or Y. enterocolitica for 6 h, change in the gene expression was evaluated through quantitative real-time PCR. The results from 3 independent experiments are presented as a fold regulation which is the negative inverse of the fold change if the fold change is less than 1.
N.S., not significant.
**p<0.01 and ***p<0.001 indicate significant differences between the groups compared.

Figure 3
Y. enterocolitica induces the activation of PKA in PP M cells. The PPs were prepared from the mice orally administered with or without CFSE-labeled Y. enterocolitica, stained with anti-GP2 Ab and anti-p-PKA substrate Ab, and counter stained with DAPI. The samples were analyzed by confocal laser scanning microscopy. The image is a stack of XY. The arrows indicate M cells. Scale bars represent 20 μm.
C5aR is critically involved in regulating Y. entercolitica infection into M cells
It is well-recognized that PKA activation mediated the weak iNOS-dependent killing of pathogens, although it was poorly studied in M cells (19). We, therefore, tried to confirm whether PKA activation in M cells has an influence on bacterial infection in M cells (Fig. 4). When we measured the survival rate of internalized Y. enterocolitica in M-like cells stimulated with or without C5a or C5aRA, C5a treatment on M-like cells during the Y. enterocolitica infection significantly (p<0.001) enhanced the survival of internalized Y. enterocolitica such that the number of survived Y. enterocolitica was about 2-fold increased by C5a treatment while blockade of C5aR activation by C5aRA treatment significantly (p<0.01) reduced the survival of internalized Y. enterocolitica. Importantly, this C5a activation-dependent increase of internalized Y. enterocolitica survival and C5aRA-depedent blockade of increased survival of internalized Y. enterocolitica survival in M-like cells was not detected in E-like cells which do not express C5aR (Fig. 4A).
Figure 4
Y. enterocolitica exploits C5aR signaling for their survival in vivo. (A) The human M-like cells and E-like cells were either pretreated or not with C5aRA and then infected with Y. enterocolitica. (B, C) The WT mice and C5aR KO mice were orally administered with Y. enterocolitica. The M-like cells (A), PP cells (B), or MLN cells (C) were prepared 3 h after infection and used to determine viable Y. enterocolitica CFU. Data are represented as mean±SE (n=3) in the group.
CFU, colony-forming unit; SE, standard error; ND, not determined.
*p<0.05, **p<0.01, and ***p<0.001 indicate significant differences between the groups compared.

We further confirmed the in vivo role of crosstalk between C5aR and TLRs in Y. enterocolitica infection into M cells by using C5aR KO mice (Fig. 4B and 4C). Three hours after oral administration of Y. enterocolitica into WT mice or C5aR KO mice, PPs and MLNs were prepared and the number of Y. enterocolitica in the tissues was measured by incubating the tissue homogenates in CIN agar plate. Consistent with the in vitro results using M-like cells shown in Fig. 4A, PPs and MLNs of C5aR KO mice contained significantly (p<0.05) reduced about 95% invaded Y. enterocolitica compared to those in WT mice. We believe that this reduction of initial infection was closely related with the bacterial clearance in M cells in the absence of C5aR-mediated signal. Collectively, these results suggest that C5aR signaling on M cells could regulate the infection of Y. enterocolitica through exploiting the crosstalk with TLR-mediated signaling.
DISCUSSION
It was reported recently that allograft inflammatory factor 1 regulates the transcytosis in M cells and affects translocation of invaded Y. enterocolitica via activation of β1 integrin (20). Our previous study also reported that C5aR is expressed in apical area of M cells and C5aR is closely associated with the Y. enterocolitica infection in M cells (8). However, although it is known that many mucosal pathogens exploit M cells for their infection, we do not clearly understand how bacterial pathogens invade into tissue by infecting M cells. In that sense, this study results suggest the mechanism for survival of mucosal pathogens in M cells because it is conceivable that inhibition of signal crosstalk between C5aR and TLRs in M cells gives an opportunity to pathogens to survive in early anti-microbial immune response. Our speculation was supported by decreased survival rate of Y. enterocolitica in human M-like cells by C5aRA treatment and by inhibited infection rate of Y. enterocolitica in C5aR KO mice (Fig. 4).
C5aR is a seven-transmembrane protein categorized in peptide receptor subfamily of class A rhodopsin-like receptors and its signaling activation involves intracellular calcium mobilization (21). The classical ligand of C5aR is C5a which is a pro-inflammatory peptide, but some bacteria-derived proteins including seventeen kilodalton protein (Skp) of gram-negative bacteria can also directly bind to C5aR (22). Except for the signal crosstalk with TLRs, C5aR can also crosstalk with IgG Fc receptors which enhance the FcγRIII-triggered cellular activation (23). Therefore, several mucosal pathogens could exploit signal crosstalk between C5aR and other receptors to modulate host functions.
Pou2af1 gene encodes POU domain class 2-associating factor 1 (POU2AF1) and its expression was enhanced in human M-like cells stimulated with Y. enterocolitica (Fig. 2) (24). Although this gene was previously characterized to be expressed specifically in lymphocytes, recent study reported its expression in human airway epithelium (25). Interestingly, POU2AF1 can directly regulate SPIB which is master transcription factor for differentiation of M cells from Lgr5+ stem cells (2627). Therefore, we expect that crosstalk between C5aR and TLRs may be also involved in M cell differentiation. Above all, this study has shown that expression of TLRs were observed in M cells expressing C5aR and cAMP-PKA pathway in M cells can be stimulated by Y. enterocolitica infection. In addition, Y. enterocolitica infection into M cells was enhanced by C5a treatment and this enhancement was abrogated by C5a antagonist treatment. Finally, Y. enterocolitica infection into M cells was unsuccessful in C5aR KO mice. Collectively, we suggest that exploit the crosstalk between C5aR and TLR signaling is one of infection mechanisms utilized by mucosal pathogens to infect M cells.
 XML Download
XML Download