

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The interaction of T cell receptor (TCR) on T cells and self-peptide (self-p) bound to major histocompatibility complex (MHC; self-pMHC) ligands on antigen-presenting cells (APCs) has a huge impact on T cell development and tolerance establishment in the thymus (12). Developing immature thymocytes die by neglect or undergo stringent apoptotic death by negative selection when they do not properly interact with or make too much strong interactions with self-pMHC ligands, respectively, and those only interacting with ‘Goldilocks’ (not too weak or not too strong) strength of self-pMHC survive (by positive selection) and go through further developmental process to enter mature naïve T cell pool (3456). The mechanisms of positive and negative selection during thymic development, along with essential cellular and molecular components, have been reviewed in details elsewhere (789) and thus will not be discussed in this review.
After successful thymic selection, mature naïve T cells enter the periphery and circulate various secondary lymphoid organs where they reside and continue to interact with a vast array of diverse self-pMHC ligands displayed on the surface of various cell types including APCs unless there is episode of invading pathogens. Over decades of studies by many investigators, it is now clear that the tonic TCR signals from peripheral self-recognition of naïve T cells is critical for their survival via a mechanism that has not been clearly identified yet. Thus, continuous TCR contacts with self-pMHC ligands, together with IL-7, is the fundamental life-sustaining event for whole life-time of peripheral naïve T cells (101112). In addition, the concept of life-long TCR self-recognition has also attracted and been extended to define its role in other important biological aspects of resting naïve T cells, particularly their functional properties in generating rapid and robust antigen-specific immune responses (131415).
The role of self-recognition in T cell function is of great importance especially when considering that upon pathogenic infections, self-pMHC ligands would far exceed in numbers over only a few cognate foreign pMHC ligands on APCs that are detectable by extremely rare precursor frequency of antigen-specific naïve T cells in vivo. Recent advances in sensitive analytic tools for detecting rare antigen-specific naïve T cell clones and their biological responses, biochemical methods for identifying endogenous self-peptides, and genetic tools for generating TCR transgenic animals have all greatly contributed to uncovering an essential feature of TCR-self-pMHC interactions and its physiological importance as a functional modulator, which has led to several central questions. For example, how do naïve T cells take advantage of their self-recognition for enhancing their functional responsiveness to antigenic stimulation? Does the strength of self-reactivity that may differ among individual naïve T cells have different impact on quantity and quality of T cell functional responsiveness, and which cell types and what factors mainly contribute to modulating the strength of self-reactivity? Is this effect by self-recognition set intrinsically for T cells during thymic development, or acquired later dynamically during their peripheral residency? Do the TCR signals from self-recognition have any regulatory influence on functional activity of naïve T cells which is required for avoiding autoimmunity?
In this brief review, we discuss the requirement of self-recognition of naïve T cells with a particular emphasis on its beneficial impact on the functional immune responses to antigenic stimulation. We also discuss recent findings on some of key factors and relevant mechanisms found thus far to link the relationship between the strength of self-reactivity and its impact on intrinsic functional activity of naïve T cells. Finally, we will end with a model of how naïve T cells utilize their ‘sense of self’ not only to maintain full functional integrity but also to shape a diverse range of functional heterogeneity distinctly different from one another.
IMPACT OF TCR RECOGNITION OF ENDOGENOUS SELF-pMHC LIGANDS ON NAÏVE T CELL RESPONSIVENESS TO COGNATE FOREIGN ANTIGENS
Effect of self-deprivation on T cell functional responsiveness to antigens
Peripheral naïve T cells continuously scan the surface of various cell types in the secondary lymphoid organs to search for their cognate foreign pMHC ligands, at the event of which leads to their activation, population expansion, and differentiation into effector cells that can eliminate foreign antigens. However, under steady state without any infectious episode, naïve T cells would not see any of cognate foreign antigens but rather have much more frequent chances to see vast arrays of endogenous self-antigens — presumably most but not all the same as those of mediating positive selection in the thymus — displayed during their peripheral residency. It is therefore of importance to determine whether, besides its role in maintaining naïve T cell survival, their TCR engagement of self-antigens may help some other biological aspects of peripheral naïve T cells.
Seminal study by Stefanová et al. (13) first demonstrated previously unidentified importance of endogenous self-recognition by TCR; that is, required for optimal T cell reactivity to foreign antigens. Using transgenic CD4+ T cells expressing MHC class II (MHC II)-restricted monoclonal TCR transgene, they revealed two very important findings. First, when naïve T cells were deprived from contacts with self-pMHC ligands (either by culturing cells in suspension to abolish TCR-self-pMHC contacts in vitro, injection of anti-MHC II antibody in vivo, or adoptive transfer into MHC II-deficient or non-selecting MHC II-expressing recipients), those T cells recovered lost their responsiveness to cognate foreign antigens (as assessed by phosphorylation of TCRξ-chain and ZAP-70, proliferative response, and IL-2 production). Second, mechanistically, functional failure of these self-deprived cells was associated with loss of polarized TCR (and ZAP-70) distribution on the membrane, an event that is required to facilitate TCR clustering and amplify signal transduction for activation upon antigen encounter.
In line with these findings, other study also demonstrated an effect of self-deprivation on CD4+ T cell responses to antigenic stimulation (16) and came to the same conclusion of the importance of TCR self-recognition but with a mechanism different from those observed by Stefanová et al. (13). In this study, OT-II TCR transgenic CD4+ T cells (specific for chicken ovalbumin [OVA] peptide 323–339 bound to MHC II I-Ab complex) deprived of self-pMHC contacts by transferring into recipient mice lacking MHC II expression failed to respond to antigenic stimulation with in vivo injection of OVA-expressing dendritic cells (DCs) (as assessed by reduced proliferation and levels of early activation marker CD69 expression). In search for a mechanism underlying this phenomenon, these authors found that the impaired response was not due to defective TCR signaling but rather reduced in vivo motility of self-deprived OT-II cells and accordingly defect in their ability to interact and make clusters with antigen-bearing DCs, which was associated with impaired cell adhesion and cytoskeletal reorganization as a result of reduced levels of small GTPases Rap1 and Rac1.
Despite seemingly different mechanisms being suggested, the above studies pointed that self-deprivation of naïve T cells for a certain period of time (several days or weeks) and even for up to 15–30 min seemed sufficient to cause the loss of antigen responsiveness (1316). It is therefore clear that peripheral naïve T cells must see self-pMHC ligands all the time to maintain adequate levels of functional ‘fitness’, thereby generating optimal antigen-specific responses.
Role of DCs as a major ‘self-presenter’ for sensitizing T cell antigen responsiveness
If the optimal T cell responses depend on continuous TCR contact with self-pMHC ligands, does all cell types or a certain subset of APCs that express relevant MHC molecules (MHC class I [MHC I] for CD8+ T cells and MHC II for CD4+ T cells) retain this unique property for optimizing T cell antigen responsiveness?
An example of a particular subset of APCs, that is, DCs, has recently been examined for this specialized role in vivo, by using mice that are transgenically engineered for removing DCs in an inducible manner (17). In this experiment, both CD4+ and CD8+ T cells purified from monoclonal TCR transgenic or polyclonal B6 mice that had been depleted of DCs for 2 days exhibited severely diminished proliferative responses to cognate foreign antigens (for monoclonal TCR transgenic T cells) as well as bacterial super antigen (staphylococcal enterotoxin A for polyclonal T cells). Notably, this impaired T cell response was reversible however; thus, the observed defect was completely restored when depleted DC counts were recovered back to normal levels or when T cells from DC-depleted mice were pre-incubated for only a short time of 30 min with DCs. Mechanistically, the impaired antigen responsiveness was associated with defect in early TCR signaling as well as in their ability to form a productive immunological synapse on the membrane interface between DCs-depleted T cells and APCs. This phenomenon seems in close agreement with the above findings of Stefanová et al. (13) demonstrating defective TCR polarization on self-pMHC-deprived naïve CD4+ T cells and also highlights the previously identified specialized property of DCs in facilitating T cell-DC synapses in an antigen-independent manner that can lead to some levels of T cell responses such as Ca2+ increase (18), although the latter effect was curiously independent of MHC expression on DCs.
Given the significant expression levels of relevant MHC molecules on cell types other than DCs in lymphoid tissues, particularly B cells or macrophages, as well as non-immune stromal cells including fibroblastic reticular cells or lymphatic endothelial cells, it will be interesting to determine whether these non-DC cell types would also have any as-yet-undefined accessary role as an authentic ‘self-presenter’ for sensitizing optimal T cell responsiveness. More importantly, what factors make DCs to exhibit such a unique property specialized for this purpose of T cell functional readiness — an issue of which is also pertinent to the more generalized but seemingly opposing effect of steady-state DCs on T cell tolerance induction (see below) — is also an important question awaiting further studies.
Role of self-ligands as a ‘coagonist’ for T cell reactivity to foreign antigens
The observed role of self-recognition described above relies on continuous interaction between peripheral naïve T cells and APCs, especially DCs, under steady state condition, and highlights that such uninterrupted self-derived tonic TCR signaling is essential for ensuring functional preparedness of T cell sensitivity and reactivity in response to foreign antigen encounter. It should be noted, however, that the above role of ‘pre-engagement’ with self-ligands does not necessarily mean that endogenous self-pMHC ligands is dispensable with negligible impact on T cell activation at the time of TCR recognition to cognate foreign antigens. Indeed, number of studies have provided compelling evidence supporting the critical requirement of simultaneous engagement of TCR for both self-pMHC ligands and foreign agonist pMHC ligands for efficient T cell activation (1920212223).
One of these studies, using 5C.C7 TCR transgenic CD4+ T cells specific for moth cytochrome c (MCC) peptide bound to MHC II I-Ek complex molecule, have shown that several endogenous self-peptides identified accumulated at the T cell-APC synapse and that soluble pMHC heterodimers — one loaded for endogenous self-peptide and the other for agonist MCC peptide — but not monomeric MHC of either self or agonist peptide, could activate naïve T cells, including a Ca2+ response and IL-2 production (20). As a mechanistic basis, this study suggested a ‘pseudodimer’ model as a basic unit required for naïve T cell activation, in which two TCRs, one of which bound to an antigenic pMHC complex and the other to an endogenous self-pMHC complex, are stabilized by CD4 co-receptor-mediated bridging (20). Two other consecutive studies using MHC II-restricted monoclonal CD4+ T cells have also found that only a specific, but not all, endogenous self-peptides that were identified to mediate most efficient positive selection in the thymus could serve as a ‘coagonist’ in conjunction with agonist antigenic peptides to augment T cell responses in terms of CD69 upregulation, proliferation, and cytokine production (2223).
Similar coagonist activities of endogenous self-peptides have also been demonstrated for activation of MHC I-restricted naïve CD8 T cells, although its underlying mechanism of action seemed distinctly different from those proposed for naïve CD4+ T cell activation (1921). All these findings from the above studies fortify a mechanistic role of self-pMHC ligands with coagonist activity other than ‘pre-conditioning’ effect on proximal TCR signaling per se. Nevertheless, the main conclusion, together with those obtained from the above self-deprivation studies, is not necessarily mutually exclusive to but rather fits very well to the hallmark features of naïve T cell responses, that is, remarkable antigen sensitivity, a response that is triggered by even a single antigenic pMHC complex molecule displayed on the surface of APCs (24).
INFLUENCE OF THE STRENGTH OF TCR SELF-RECOGNITION ON THEIR ANTIGEN-SPECIFIC IMMUNE RESPONSES
Peripheral naïve T cell populations are expected to display variable strength of self-reactivity, because their thymic precursor cells receive a weak but diverse levels of TCR signals for positive selection within the permissive range of TCR affinity spectrum for self-pMHC ligands. Thus, in light of the above established role of self, it is tempting to speculate that the difference in the strength of TCR self-recognition would result in a somewhat distinct functional outcome different from one another among peripheral naïve T cell pool. This issue, however, seems difficult to address by virtue of extremely low TCR affinity for binding to endogenous self-pMHC ligands which is impractical to measure its signaling response with current biochemical methods. Instead, in an attempt for tracking diverse strength of TCR self-reactivity, it has been well documented that intrinsic TCR affinity for self-pMHC ligands correlates with expression levels of T cell surface protein CD5, a putative negative regulator for TCR signaling, whose levels are developmentally set during thymic positive selection (252627).
Using this correlation between TCR affinity for self-ligands and CD5 levels, effects of variable strength of TCR self-recognition on naïve T cell function have recently been addressed by several research groups. In this regard, our previous study (28) revealed a marked functional difference in T cell responses to cytokines, including IL-2 and IL-15, with polyclonal (and also monoclonal TCR transgenic) naïve CD8+ T cells expressing high levels of CD5 (CD5hi) being greater proliferative responses than those of naïve CD8+ T cells expressing low levels of CD5 (CD5lo). We found that this phenomenon was a result of enhanced cytokine receptor clustering and reorganization upon ligand engagement through higher levels of membrane lipid rafts on CD5hi cells than CD5lo cells, therefore providing selective advantage to CD5hi cells over CD5lo cells and consequently resulting in better proliferative responses of the former subpopulation to relatively a weak antigen in vitro.
Similar to our in vitro findings, other recent two studies using polyclonal CD4+ or CD8+ T cell populations along with the in vivo infection models have also observed a superior functional outcome of CD5hi cells relative to their CD5lo counterparts in terms of antigen-specific activation and expansion responses after infection with viral or bacterial pathogens (1415). Despite the same conclusion being drawn from these two observations, however, each of these studies suggested distinctly different mechanisms behind this phenomenon. The results of Mandl et al. (for naïve CD4+ T cells) (14) provided several compelling evidences supporting seemingly intriguing possibility that thymic positive selection drives preferential skewing mature TCR repertoire toward higher affinity populations for both self and foreign antigens; thus, CD4+ T cell self-reactivity was positively correlated with TCR strength for cognate foreign antigens, resulting in greater antigen-specific expansion of CD5hi cells than CD5lo cells. By contrast, the data for naïve CD8+ T cells from Fulton et al. (15) indicated, however, that although similar in the functional outcome of CD5hi cells being dominated with greater antigen-specific responses than those of CD5lo cells, this phenomenon was not a result of different binding affinity to foreign pMHC ligands but rather associated with better functional competence of the former subpopulation, for instance its effector/memory-associated gene signatures and higher sensitivity to extrinsic stimulatory and/or inflammatory cues.
In close accordance with this latter phenomenon, recent report by Persaud et al. (29) also revealed that, using two distinct monoclonal TCR transgenic CD4+ T cells that are specific for an identical foreign antigen but differ in their levels of CD5 expression, CD5hi cells exhibited a greater response for IL-2 production and ERK phosphorylation than CD5lo cells following stimulation with specific antigenic peptides, antibody to the invariant signaling protein CD3 (anti-CD3) plus antibody to the costimulatory molecule CD28 (anti-CD28), or phorbol 12-myristate 13-acetate (PMA) plus ionomycin. These findings were also confirmed for polyclonal CD4+ and CD8+ T cell populations and even for single-positive (SP) but not immature double-positive (DP) thymocytes, and showed that the enhanced functions of CD5hi subpopulation were lost when the cells were deprived of self-pMHC contacts for several days (29). These data therefore suggest that, together with the data obtained from short-term MHC deprivation in vivo, the observed superior functional property of higher self-reactive T cell populations (i.e., CD5hi cells) might be set during thymic positive selection and persists with the continuous requirement of peripheral TCR self-recognition.
A common theme from all these studies described above centers on the fundamental and conceptual issue of the major purpose of thymic positive selection; that is, shaping functional heterogeneity of mature T cell pool with some subpopulations being better poised to drive effective immune responses than the others. There are, however, several important questions being open. For example, precisely how such ‘coincidence’ of selecting ‘good’ TCRs for both self- and foreign-ligands (as reported by Mandl et al.) (14) is regulated during thymic selection process, and how the ‘pre-conditioning’ of higher self-reactive T cells (as illustrated by Fulton et al. and Persaud et al.) (1529) is linked to ‘wiring’ them to acquire better functional competence remains largely unclear and warrants interesting new subjects for future studies.
REGULATORY IMPACT OF TCR SELF-RECOGNITION ON T CELL RESPONSIVENESS
While peripheral naïve T cells require continuous TCR-self-pMHC interactions for maintaining functional competence to exert optimal responses following subsequent antigenic stimulation, these populations under steady state must also be in check to be quiescent in their activity without inducing overt responses to any self-components to avoid harmful autoimmunity. How can possibly individual TCR signals from contacts with self-pMHC ligands deliver a highly-controlled form of such distinct and seemingly opposing effects on naïve T cells, that is, functional enhancement to foreign antigens vs. dampening reactivity to self-ligands? Indeed, there are number of evidences supporting the role of TCR-self interactions in ‘tuning’ T cell antigen reactivity via modulating its activation threshold with several different mechanisms, including dynamic regulation of either surface or intracellular levels of negative (or positive) regulatory proteins, their subcellular distribution or localization, and transcriptional regulation via microRNA (303132333435). Such tuning by TCR-self interaction seems to occur at two different stages: one for developing thymocytes in the thymus (called ‘developmental tuning’) and the other for resting naive T cells in peripheral lymphoid organs (called ‘peripheral tuning’).
Developmental tuning
It has long been known that preselection thymocytes undergoing positive selection are subjected to TCR tuning by virtue of controlled expression of CD5, a negative regulator for TCR signaling, whose expression on thymocytes is induced as a consequence of TCR interactions with positively selecting self-pMHC ligands (252627). Since the levels of CD5 are dynamically regulated and proportional to the strength of TCR-self interactions, thymocytes with higher self-reactive TCR would induce enhanced levels of CD5 for the adaptation to the strong TCR signaling via increasing activation threshold for negative selection, thereby directing them to be positively selected, which otherwise being deleted. Therefore, TCR self-recognition utilizes dynamic regulation of CD5 expression for playing a pivotal role in fine-tuning TCR signaling and broadening the affinity spectrum of positive selection of TCR repertoire. Number of studies have indeed supported this notion of ‘developmental tuning’, including the results showing that thymocytes from CD5-deficient mice were hyper-responsive to TCR engagement (25), presumably via lowering the threshold for TCR activation, and moreover, that mature TCR repertoire was largely altered in mutant mice either lacking (253637) or overexpressing CD5 proteins (27).
Similar to the tuning effect with CD5 on developing thymocytes, it should be noted that other surface protein CD6 was also illustrated to have a negative effect on TCR signaling, presumably involving developmental tuning for high self-reactive TCR during thymocyte development, as evidenced by enhanced Ca2+ flux responses of thymocytes from CD6-deficient mice (38); given its association with CD5 on the T cell surface (39), however, whether this effect by CD6 deficiency may be secondary to modulating CD5 expression remains to be determined. In any case of either CD5 or CD6, these findings strongly suggest the importance of developmental tuning during selection stages in the thymus, which is required not only for dampening excessive TCR signals from interactions with high affinity self-pMHC ligands — a strength of which is around interface between positive and negative selection — but also for generating a diverse range of TCR repertoire with much broader but permissive self-reactivity.
Peripheral tuning
Besides the above developmental tuning, studies have also provided evidences supporting the existence of such tuning for steady-state resting naïve T cell populations (35). As for the former occurring in the thymus, the latter ‘peripheral tuning’ is also mediated through a mechanism involving continuous TCR interactions with self-pMHC ligands in peripheral lymphoid tissues. In support of this, results by Smith et al. (40) indicated that monoclonal TCR transgenic naïve CD4+ T cells (from AND TCR mice) displayed enhanced Ca2+ responses to TCR stimulation with anti-CD3 or cognate antigenic peptides when these cells were deprived of MHC II contacts for 3 days in a 3-dimensional reaggregate culture. Because this phenomenon seemed associated with down-modulation of CD5 expression on the MHC-deprived AND T cells, they explored then to compare CD5lo and CD5hi fractions of polyclonal naïve CD4+ T cell populations and came to a surprising observation showing the greater anti-CD3-induced Ca2+ responses for CD5lo cells than those for CD5hi cells (40), suggesting an obvious inverse correlation of CD5 expression levels and, at least for CD4+ T cells, their antigen responsiveness.
In vivo physiological relevance of such peripheral tuning came from a study by Bhandoola et al. (41), in which CD4+ T cells were adoptively transferred into either MHC II-sufficient or MHC II-deficient recipient mice on a lymphopenic background (lacking ZAP-70). In this study, the donor CD4+ T cells in both recipients showed similar levels of population expansion over a month of time period with acquisition of activated memory phenotypes (CD44hi CD62Llo), but surprisingly, resulted in a quite different outcome in responses to syngeneic MHC II+ skin grafts; while mice receiving CD4+ cells in MHC II-sufficient environment accepted the grafts, those receiving CD4+ cells in MHC II-deficient environment rejected the grafts with exaggerated intraepithelial infiltrations of donor CD4+ cells (41). These results suggest that peripheral self-MHC II expression and its continuous interactions with naïve CD4+ T cells might be important to restrain their TCR auto-reactivity and maintain peripheral self-tolerance, presumably via two complementary mechanisms, including direct TCR tuning as well as suppression by CD4+ T regulatory cells maintained in the presence of self-MHC II.
Likewise, the peripheral tuning through TCR-self interactions was also illustrated for steady-state resting naïve CD8+ T cell population. Recent study by Takada and Jameson (42) demonstrated that using a non-lymphopenic setting, MHC I deprivation (for about 14 days) led to an augmentation of naïve CD8+ T cell responses to a weaker, but not strong, antigenic peptides (as assessed by upregulation of early activation marker CD69). In this study, however, the enhanced response of MHC-deprived cells was not associated with CD5 expression levels, but instead proposed to occur as a result of upregulated expression of CD8 co-receptor, whose transcription was found to be negatively regulated by tonic TCR signals from its self-recognition (42). Recently, we also addressed this issue of peripheral TCR tuning by comparing CD5lo and CD5hi cells of resting naïve CD8+ T cell populations and determining whether higher self-reactive T cells are indeed more ‘tuned’ in their TCR signaling sensitivity, leading to a reduced responsiveness to subsequent TCR engagement (43). The results from our experiment showed that CD5hi cells did exhibit diminished TCR responses compared to those of CD5lo cells, as assessed by reduction in Ca2+ response and in phosphorylation of proximal TCR signaling components (ERK, ZAP-70, and PLC-γ), as well as decrease in upregulation of CD69 and in downregulation of CD62L (38). We also found that the TCR tuning that is predominant for CD5hi naïve CD8 T cells correlated with their lower basal levels of active p56LCK and higher surface expression of CD45, a membrane tyrosine phosphatase that regulates p56LCK activity (43).
Despite these supportive evidences, it is of importance to note that the tuning effect is apparently in contrast to the positive role of TCR-self interactions required in general for optimal T cell responses as described in the prior sections of this review. Nevertheless, all results from the above studies including ours highlight a key notion that, both for naïve CD4+ and CD8+ T cells, continuous TCR self-recognition (especially those for higher self-reactive cells) would attenuate TCR sensitivity through dynamic tuning of surface levels of several different negative and/or positive regulatory proteins, including CD2 (44), CD5 (40), CD6 (38), CD8 (42), and CD45 (43), as well as, although not discussed here in detail, expression of intracellular signaling proteins, whose functions negatively regulate TCR signaling, such as various protein tyrosine phosphatases, particularly, PTPN2 and PTPN22 (3334).
Despite the close functional similarity akin to those of developmental tuning, the peripheral tuning seems to have a more unique and directed effect on maintaining self-tolerance of high self-reactive T cell populations. It is therefore interesting to determine whether DCs contribute as a major cell type of providing a unique regulatory impact on promoting peripheral tuning process for naïve T cells. In this regard, several studies have shown that steady-state DCs have unique capacity to induce a strong form of tolerance for T cell responses to a model self-antigen or even cognate foreign antigens (4546474849). Likewise, other non-immune cell types such as stromal cells residing in peripheral lymphoid tissues have also been reported for their unique tolerance-inducing properties to peripheral tissue self-antigens (505152). It should be noted, however, that the result of the tolerance mediated by these cell types is expected to become either of two outcomes: that is, deletion or functional unresponsiveness in an irreversible manner. Therefore, regardless of no matter what cell types are involved, the peripheral tuning for naïve T cell populations must be precisely controlled to fine-tune TCR sensitivity just enough to induce survival signals but not activation signals to self-ligands, without causing destructive tolerance of their functional integrity.
CONCLUDING REMARKS
TCR self-recognition has a huge impact on almost entire period of T cell's life-span. On the one hand, it is required for the selection and development of mature T cell repertoire in the thymus as well as for the maintenance of their survival and acquisition of full functional competence during steady-state peripheral residency. In particular, the latter properties of T cells, called ‘self-conditioning’ here, are critical for the induction of rapid and robust T cell immune responses to subsequent antigenic encounter. On the other hand, the signals from TCR-self interactions are equally important, however, for the attenuation of excessive TCR reactivity to self-components, a phenomenon of ‘self-tuning’ that is required for avoiding autoimmunity. Therefore, tonic TCR signals derived from self-recognition must be tightly regulated all the time but also have some degree of flexibility, which depends on the relative intensity of self-reactivity of individual TCRs, to exert such dual functionalities on T cell homeostasis.
Although much progress has been made to understand the positive and negative impact of TCR self-recognition on T cell functional responses, several unresolved questions remain. How do TCR-self interactions regulate the functional fate of either conditioning or tuning of diverse naïve T cell repertoire? Are these processes regulated in distinctly different manner or are they shared in balance at some points? What factors (or environmental contexts) determine which processes are dominated? If the two different functional outcomes (conditioning vs. tuning) share some common pathways that are not mutually exclusive and their nature and balance are probably determined by the strength of self-TCR signals, it is then possible that the seemingly opposing two functional outputs described above may be dominant for higher self-reactive T cells, as illustrated in Fig. 1. We thus propose that TCR's ‘sense of self’ is of particular importance for selecting those of potentially auto-reactive T cell clones in the thymus and adding them into steady-state normal T cell repertoire in the periphery, where they continue to be tailored to be quiescent in function while being also nurtured for keeping full functional fitness ready for inducing rapid and robust immunity upon pathogen infections.
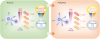 | Figure 1Schematic diagram illustrating impact of TCR self-reactivity on shaping phenotypic and functional heterogeneity of peripheral naïve T cells. Both in thymus and periphery, self-recognitions by TCR heavily affect CD5 levels on T cells correlating with the strength of TCR affinity for self-ligands (i.e., low, intermediate, and high levels of CD5; CD5lo, CD5int, and CD5hi, respectively). Here the CD5hi cells relative to CD5lo cells are likely to be more ‘tuned’ both in the thymus and in the periphery, but at the same time better ‘conditioned’ in their functionality in response to various homeostatic or stimulatory and inflammatory cues, which allows in turn for these high self-reactive cells not only to generate more heterogeneity in their phenotypes and also to shape their function in dominating anti-viral immune responses.
CD5int, intermediate level of CD5.
|
 XML Download
XML Download