

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The ultimate goal of periodontal treatment is to prevent further loss of attachment and to regenerate periodontal supporting tissues including cementum, periodontal ligament (PDL), and bone. To achieve periodontal regeneration, several therapeutic approaches have been examined in animal models and clinical studies, including bone grafting, guided tissue regeneration, and the implantation of biologic mediators [1]. However, inconsistent beneficial effects of these treatments have been observed. The application of magnetic fields has been used as an alternative and/or complementary treatment for fracture healing, spinal fusion, osteoarthritis, and wound healing [2]. Magnetotherapy including static magnetic fields (SMFs), pulsed electromagnetic fields (PEMFs), and alternating electromagnetic fields provides a non-invasive, safe, and easy method to directly treat the site of injury, the source of pain and inflammation, and various types of disease. Although PEMFs have been used extensively in clinical treatments of non-united bone fractures for a long time [3], there is some skepticism concerning the proposed exposure protocols [4]. Furthermore, PEMFs are generated by electrical devices.
SMFs are a type of magnetic field used in clinical practice, especially in the dental field [5], because SMFs do not need power devices, as PEMFs do. In addition, SMFs are convenient and suitable for long-term application [6], and do not provide any heat or electric hazard to surrounding tissue [7]. Rare-earth magnets, which generate SMFs, have been used in magnetic retention appliances in implant- or tooth-retained overdentures [8], maxillofacial prostheses after trauma and cancer surgery [9], as well as in orthodontic treatments such as space closure, molar distalization, intrusion, the traction of impacted teeth, and palatal expansion [5]. SMFs increased implant stability and reduced bone loss during the initial weeks of healing [10]. Recently, we demonstrated that exposing sandblasted, large-grit, acid-etched-treated titanium implants to 15-mT SMFs triggered faster early peri-implant bone formation in rabbits [11].
However, the effect of SMFs on growth and osteoblastic differentiation remains controversial. SMF exposure had no significant effect on the proliferation of rat calvaria cells or osteoblast-like cells [12]. The proliferation of human osteoblast cultures was reduced when cells were exposed to continuous low-intensity SMFs [13]. Moreover, SMFs generated by corrosion currents inhibited human osteoblast differentiation [14]. In contrast, SMFs were found to increase the osteoblastic differentiation of rat osteoblasts [12], human osteoblast-like MG63 cells [18], and dental pulp cells [16]. In addition, we found that 15-mT SMFs, an intensity that is relatively low within the intensity range of moderate SMFs (1 mT to 1 T), enhanced the proliferation and osteogenic differentiation of human bone marrow-derived mesenchymal stem cells (MSCs) in vitro [17].
Periodontal regeneration involves the recruitment of locally-derived progenitor cells that can subsequently differentiate into periodontal ligament cells (PDLCs), mineral-forming cementoblasts, or bone-forming osteoblasts [18]. Although SMFs can enhance bone fracture healing and bone formation by osteoblasts both in vivo and in vitro [1920], their effects on the differentiation of PDLCs, cementoblasts, and osteoblasts are not fully understood. These cells constitute the periodontium and can be directly influenced when SMFs are applied to teeth, implants, and other intraoral structures. Thus, the aim of this study was to investigate the effects of SMFs (15 mT) on the osteoblastic and cementoblastic differentiation of human PDLCs, cementoblasts, and osteoblasts, and to explore the possible mechanisms underlying these effects.
MATERIALS AND METHODS
Cell culture
Human fetal osteoblasts (hFOB 1.19) were obtained from the American Type Culture Collection (CRL-11372, Rockville, MD, USA) and maintained in Dulbecco's modified Eagle's medium-Ham's F-12 (1:1) basal medium supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin/glutamine (Gibco, 15140-122, Invitrogen Corporation, Grand Island, NY, USA) in a standard cell-culture incubator at 37°C and 5% CO2. Differentiation was induced by transfer to osteogenic medium (OM) containing 100 µM ascorbic acid, 2 mM β-glycerophosphate, and 10−7 M dexamethasone, as previously described [21].
Human PDLCs and cementoblasts [22], immortalized by transfection with the telomerase catalytic subunit of the human telomerase reverse transcriptase, were kindly provided by Professor Takashi Takata (Hiroshima University, Hiroshima, Japan). Cells were cultured in α-minimum essential medium (α-MEM; Invitrogen Corporation) supplemented with 10% FBS, 100 U/mL of penicillin, and 100 μg/mL of streptomycin in a humidified atmosphere of 5% CO2 at 37°C.
Primary osteoblasts were enzymatically isolated from the calvaria of 1-day-old ICR mice (Samtako, Osan, Korea). Briefly, the calvaria were aseptically dissected, and the fragments were incubated in phosphate-buffered saline (PBS) solution with 0.2% collagenase-dispase solution (Sigma-Aldrich, St. Louis, MO, USA) for 2 hours at 37°C. The cells were dissociated from the bone fragments, counted, and plated at 4×104 cells/cm2. The cells were grown in α-MEM supplemented with 10% FBS, 100 U/mL of penicillin, and 100 μg/mL of streptomycin in a humidified atmosphere of 5% CO2 at 37°C.
All animal experimental procedures in the current study were approved by the Institutional Animal Care and Use Committee of Kyung Hee University (approval number: KHMC-IACUC 2015-002). Primary human PDLCs were cultured using an explant technique, as described previously [23]. Briefly, impacted third molars were obtained from 3 healthy patients of the Department of Oral and Maxillofacial Surgery, Kyung Hee University Dental Hospital, Seoul, Korea. The experiments were performed with approval from the Ethical Committee of Kyung Hee University Dental Hospital, and informed written consent was obtained from all patients.
Freshly extracted teeth were immediately placed in PBS supplemented with antibiotics (100 U/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of Fungizone). In order to culture PDLCs, the PDL was carefully removed from the middle third of the root by a scalpel. Tissues were cultured in Dulbecco's modified Eagle's medium supplemented with 10% FBS and antibiotics. Cultures were maintained at 37°C in a humidified atmosphere of 5% CO2 and 95% air. Confluent cells were detached with 0.25% trypsin and 0.05% EDTA for 5 minutes, and aliquots of separated cells were subcultured. Cell cultures between the third and eighth passages were used in this study.
The SMF exposure system
A neodymium (Nd2Fe14B) disc magnet (1 mm thick and 15.0 mm in diameter, Qingdao Qiangsheng Magnets Co., Ltd., Shandong, China) and 4-well plastic culture plates were used to create an SMF exposure system (Figure 1A). A magnet was placed below the wells horizontally, and the north (N) side of the magnet was oriented towards the well. The desired field intensities (3, 15, and 50 mT) were established by controlling the distance between the magnet and the culture plates, as previously described [17]. The distance was fixed by the space between the magnet and bottom wall of the well. We measured the SMF intensity with a Gauss meter (TM-701, Kanetec, Tokyo, Japan) that could detect either axial or transverse fields. To eliminate the interference of adjacent SMFs, we attached only 1 magnet disc on 1 culture plate and provided adequate space between adjacent culture plates (at least 30 mm).
 | Figure 1Effects of SMFs on osteoblastic differentiation in human osteoblasts. (A) Schematic diagram of the SMF exposure system. (B) Cell proliferation was examined by an MTT assay at 3, 7, and 14 days. (C) Differentiation was assessed based on ALP activity, (D) Alizarin red staining, (E) calcium content, and (F) expression of the mRNA of bone matrix proteins. The intensity of Alizarin red staining was determined by optical density. Cells were treated with OS medium containing 50 μg/mL of L-ascorbic acid and 10 mM β-glycerophosphate along with 3-, 15-, and 50-mT SMFs for 7 days and 14 days. OM contained OS and 10−7 M dexamethasone. The results are representative of 5 independent experiments that were performed.
SMF: static magnetic field, MTT: 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, ALP: alkaline phosphatase, OS: osteogenic supplement, OM: osteogenic medium, Runx2: runt-related transcription factor 2, OPN: osteopontin, OCN: osteocalcin, GAPDH: glyceraldehyde 3-phosphate dehydrogenase.
a)Significant difference compared to control (P<0.05); b)Significant difference between each group (P<0.05).
|
Cell viability
Cell viability was confirmed using a 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTT) assay kit (CellTiter 96 AQueous One Solution, Promega, Madison, WI, USA) following the protocol suggested by the manufacturer.
Alkaline phosphatase (ALP) activity
Intracellular and soluble ALP, a marker of osteoblast activity, was assayed by the release of p-nitrophenol from p-nitrophenylphosphate. Cells were washed with PBS and sonicated with a cell disruptor. ALP activity was measured using p-nitrophenyl phosphate (final concentration, 3 mM) in 0.7 M 2-aminomethyl-1-propanol (pH 10.3) and 6.7 mM MgCl2. Absorbance was measured at 415 nm using an enzyme-linked immunosorbent assay reader (Beckman Coulter, Fullerton, CA, USA).
Alizarin red staining
Alizarin red staining was used to detect mineralization. Cells were fixed in 70% ice-cold ethanol for 1 hour and rinsed with distilled water. Fixed cells were stained with 40 mM Alizarin red (pH 4.2) for 10 minutes with gentle agitation. All Alizarin red staining images were photographed.
RNA isolation and reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was extracted using Trizol reagent (Life Technologies, Gaithersburg, MD, USA). Then, 1 mg of RNA was reverse transcribed using oligo (dT) 15 primers (Roche Diagnostics, Mannheim, Germany) and AccuPower RT PreMix (Bioneer Corporation, Daejeon, Korea). Thereafter, the reverse transcriptase-generated DNA (2–5 mL) was amplified. Primer sequences are detailed in Table 1. The polymerase chain reaction (PCR) products were subjected to electrophoresis on 1.5% agarose gels, and stained with ethidium bromide.
Table 1
RT-PCR primers and conditions

RT-PCR: reverse transcription polymerase chain reaction, OPN: osteopontin, OCN: osteocalcin, Runx2: runt-related transcription factor 2, CEMP-1: cementum protein 1, CAP: cementum-derived attachment protein, GAPDH: glyceraldehyde 3-phosphate dehydrogenase.
![]()
Western blot analysis
Aliquots of 1×106 cells were solubilized in ice-cold 1% Triton X-100 lysis buffer (Boston BioProducts, Inc., Ashland, MA, USA). After 30 minutes on ice, the lysates were clarified by centrifugation. Proteins (20 µg) were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (10% acrylamide), transferred to nitrocellulose membranes, and probed with specific antibodies (diluted to 1:1,000), followed by incubation with secondary horseradish peroxidase-conjugated antibody (1:5,000). Products were detected using an enhanced chemiluminescence system (Amersham-Pharmacia, Piscataway, NJ, USA).
Ca2+ assays
To measure calcium content, cells were harvested by centrifugation at 13,500 rpm for 1 minutes, resuspended in lysis buffer (0.1% Triton X-100, Boston BioProducts, Inc.) and sonicated on ice for 1 minute to destroy the cell membranes. The resulting supernatant was used for calcium measurements made with a QuantiChrom™ Calcium Assay Kit (DICA-500, BioAssay Systems, Hayward, CA, USA). Calcium content was estimated by measuring absorbance at 612 nm in a microplate reader.
Immunocytochemistry
Cells were fixed with freshly prepared 3% paraformaldehyde for 15 minutes and permeabilized with 0.5% Triton X-100 (Boston BioProducts, Inc.) for 15 minutes. After 1 hours of incubation with 10% normal goat serum/PBS, cells were incubated with anti-p65 antibody (Santa Cruz Laboratories, Santa Cruz, CA, USA) diluted 1:50 in PBS for 2 hours, washed and incubated with fluorescein isothiocyanate (FITC)-conjugated immunoglobulin G, and diluted at 1:100 in PBS for 1 hour. In order to identify the nuclei, the FITC-labeled samples were counterstained with 10 μg/mL of propidium iodide for 2 minutes. To acquire the immunofluorescence images, a confocal microscope (Cell Voyager, Yokogawa, Japan) was used.
Statistical analysis
Descriptive statistics (mean±standard deviation) for each parameter were evaluated in all the groups. The data distributions satisfied normality as tested by the Shapiro-Wilk test, meaning that intergroup comparisons could be made by using 1-way analysis of variance and the Duncan multiple range test. P values <0.05 were considered to indicate statistical significance.
RESULTS
SMFs promote osteoblastic differentiation in human osteoblasts
To determine the cytotoxic potential of SMFs, its effect on viability of the human osteoblasts was evaluated. Up to a magnitude of 50 mT, no cytotoxic effects could be observed using the MTT assay (Figure 1B). To determine the effects of SMFs on osteoblastic differentiation, we evaluated its effects on ALP activity (an early marker of osteoblastic differentiation) and mineralization. ALP activity was significantly higher (P<0.05) and calcified nodules more frequent after SMF treatment than when the cells were simply maintained in normal osteogenic supplement (OS) or OM (Figure 1C). Osteoblasts exposed to an SMF of 15 mT had significantly higher ALP levels than the other groups (P<0.05). Because the endpoint of osteoblast formation is the production of a calcified, mineralized matrix, calcium content was measured as an indicator of osteodifferentiation. The SMF-exposed osteoblasts had significantly larger mineralized nodules (Figure 1D) and extracellular calcium deposits than their control counterparts (Figure 1E) (P<0.05). The largest number of mineralized nodules and the greatest calcium content was observed in the 15 mT group. In order to explore the effects of SMFs on the osteoblastic differentiation of osteoblasts, we examined the expression of specific markers of osteoblastic differentiation. Treating the osteoblasts with SMFs increased the expression of mRNA encoding the OS- and OM-induced differentiation markers runt-related transcription factor 2 (Runx2), osteopontin (OPN), and osteocalcin (OCN) (Figure 1F and G). Maximal upregulation of the osteoblast marker genes was observed at 15 mT.
SMFs promote osteoblastic/cementoblastic differentiation in PDLCs and cementoblasts
Since SMFs of 15 mT maximally stimulated the osteoblastic differentiation of human osteoblasts, 15 mT was used in further experiments. Cytotoxicity of PDLCs and cementoblasts in response to SMFs was not observed on days 3, 7, or 14 (Figure 2A and B). To investigate whether SMFs stimulated PDLCs and cementoblasts to differentiate toward a cementoblast/osteoblast phenotype, we measured ALP activity and the expression of osteoblast- and cementum-related genes, calcium content, and calcified nodule formation. Treatment with SMFs enhanced ALP activity and mineralized nodule formation in PDLCs and cementoblasts, compared with OS and OM controls (Figure 2C-F). Similar increases in calcium concentration were also observed in SMF-treated PDLCs and cementoblasts (Figure 3A and B). In addition, SMFs upregulated the mRNA expression of osteoblast markers in PDLCs, including Runx2, OPN, and OCN (Figure 3C), as well as the expression of cementoblast markers, such as cementum protein 1 (CEMP-1) and cementum-derived attachment protein (CAP), in cementoblasts (Figure 3D).
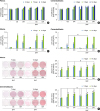 | Figure 2Effect of SMFs on osteoblastic differentiation in a cell line of human PDLCs and cementoblasts. (A, B) Cell proliferation was examined by an MTT assay at 3, 7, and 14 days. (C, D) ALP activity was determined and normalized to protein content. (E, F) Matrix mineralization was evaluated by Alizarin red staining. Cells were treated with 15-mT SMFs and OS or OM for 14 days. The results are representative of 5 independent experiments.
SMF: static magnetic field, PDLC: periodontal ligament cell, MTT: 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, ALP: alkaline phosphatase, OS: osteogenic supplement, OM: osteogenic medium.
a)Significant difference compared to control (P<0.05); b)Significant difference between each group (P<0.05).
|
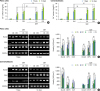 | Figure 3Effects of SMFs on calcium content and osteoblastic or cementoblastic gene expression in a cell line of human PDLCs and cementoblasts. (A, B) Ca2+ concentration was measured using a calcium quantification assay kit. (C, D) mRNA was determined by RT-PCR analysis. Cells were treated with a 15-mT SMF and OS or OM for 14 days. The results are representative of 5 independent experiments.
SMF: static magnetic field, PDL: periodontal ligament, PDLC: periodontal ligament cell, RT-PCR: reverse transcription polymerase chain reaction, OS: osteogenic supplement, OM: osteogenic medium, Runx2: runt-related transcription factor 2, OPN: osteopontin, OCN: osteocalcin, GAPDH: glyceraldehyde 3-phosphate dehydrogenase, CEMP-1: cementum protein 1, CAP: cementum-derived attachment protein.
a)Significant difference compared to control (P<0.05); b)Significant difference between each group (P<0.05).
|
SMFs promote osteoblastic differentiation in primary cultured osteoblasts and PDLCs
To further assess the osteoblastic activity of the SMFs, we examined primary cultured osteoblasts and PDLCs. Primary cultured PDLCs and osteoblasts showed no cytotoxicity in response to SMFs, but did show increased ALP activity and mineralization (Figures 4A-F, 5A and B). In addition, the treatment of primary cultured PDLCs and osteoblasts with SMFs increased the expression of mRNA encoding the differentiation markers Runx2, OPN, and OCN (Figure 5C and D).
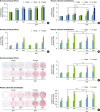 | Figure 4Effect of SMFs on proliferation, ALP activity, and mineralized nodule formation in primary cultured human PDLCs and osteoblasts. (A, B) Cell proliferation was examined by an MTT assay at 3, 7, and 14 days. (C, D) ALP activity was determined and normalized to protein content. (E, F) Matrix mineralization was evaluated by Alizarin red staining as described in the Materials and Methods. Cells were treated with 15-mT SMFs and OS or OM for 14 days. The results are representative of 5 independent experiments.
SMF: static magnetic field, ALP: alkaline phosphatase, PDLC: periodontal ligament cell, MTT: 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, OS: osteogenic supplement, OM: osteogenic medium.
a)Significant difference compared to control (P<0.05); b)Significant difference between each group (P<0.05).
|
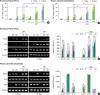 | Figure 5Effect of SMFs on calcium content and osteoblastic gene expression in primary cultured human PDLCs and osteoblasts. (A, B) Ca2+ concentration was measured using a calcium quantification assay kit. (C, D) mRNA was determined by RT-PCR analysis. The results are representative of 5 independent experiments.
SMF: static magnetic field, PDLC: periodontal ligament cell, RT-PCR: reverse transcription polymerase chain reaction, Runx2: runt-related transcription factor 2, OPN: osteopontin, OCN: osteocalcin, GAPDH: glyceraldehyde 3-phosphate dehydrogenase.
a)Significant difference compared to control (P<0.05); b)Significant difference between each group (P<0.05).
|
SMFs activate Wnt/β-catenin, p38 and c-Jun N-terminal kinase (JNK) mitogen-activated protein kinases (MAPKs), and nuclear factor-κB (NF-κB) signaling, but not extracellular signal-regulated kinase (ERK) signaling
To determine the basis of the effects of SMFs on osteoblast differentiation, their effects on Wnt/β-catenin, MAPK, and NF-κB signaling were examined. As shown in Figure 6A and B, SMFs increased Wnt1 and Wnt3a expression, the phosphorylation of glycogen synthase kinase-3β (GSK-3β), and total β-catenin protein levels in osteoblasts. Moreover, treatment with SMFs increased OM-induced p38 and JNK phosphorylation, but not that of ERK (Figure 6C). In addition, it enhanced the OM-induced nuclear translocation of p65 (Figure 6D). Immunocytochemical observations showed that the nuclear content of NF-κB p65 increased markedly in SMF-treated osteoblasts compared with the OM control (Figure 6E).
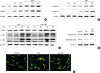 | Figure 6Effects of SMFs on the Wnt/β-catenin, MAPK, and NF-κB signaling pathways in human osteoblasts. (A, B) Cells were treated with SMFs and OS or OM for 2 days, (C) 30 minutes, and (D, E) 45 minutes. (A-D) Protein levels were assessed by western blot analysis and (E) immunofluorescence staining. Arrows (yellow) indicate the nuclear translocation of NF-κB p65. The data presented are representative of five independent experiments.
SMF: static magnetic field, MAPK: mitogen-activated protein kinase, NF-κB: nuclear factor-κB, OS: osteogenic supplement, OM: osteogenic medium, GSK-3β: glycogen synthase kinase-3β, ERK: extracellular signal-regulated kinase, JNK: c-Jun N-terminal kinase.
|
To confirm the involvement of Wnt/β-catenin, MAPKs, and NF-κB pathways in SMF-induced osteoblastic differentiation, inhibitors of Wnt (DKK1), ERK (PD98059), JNK (SP6100126), p38 (SB203580), MAPK, and NF-κB (PDTC) signaling were used. SMF-induced ALP activity, mineralization nodule formation, calcium concentration, and the mRNAs of osteoblast markers were effectively inhibited by DKK1, SP600126, SB203580, and PDTC, but not by PD98059 (Figure 7A-D).
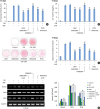 | Figure 7Effects of various inhibitors of signal transduction on SMF-induced osteoblastic differentiation in human osteoblasts. (A-D) Cells were pretreated for 2 hours with DKK1 (0.5 μg/mL), SB203580 (20 μM), PD98059 (20 μM), SP600125 (10 μM), and PDTC (10 μM), and then cultured in OS with 15-mT SMFs for 14 days. The results are representative of 5 independent experiments.
SMF: static magnetic field, OS: osteogenic supplement, Runx2: runt-related transcription factor 2, OPN: osteopontin, OCN: osteocalcin, GAPDH: glyceraldehyde 3-phosphate dehydrogenase.
a)Significant difference between each group (P<0.05).
|
DISCUSSION
Successful periodontal reconstruction requires the regeneration of multiple tissues, including cementum, PDL, and bone. Osteoblasts are specialized mesenchymal-derived cells capable of forming new bone. PDLCs retain the capability to differentiate into the osteoblast lineage, and are considered to play a key role in the regeneration of periodontal tissues [24]. In addition, cementoblasts may play a key role in the formation of the PDL-cementum apparatus during periodontal regeneration [25]. Thus, complete periodontal repair entails de novo cementogenesis, osteogenesis, and the formation of PDL fibers, and we focused here on the effects of SMFs on the osteoblastic/cementoblastic responsiveness of cells associated with the periodontium such as osteoblasts, cementoblasts, and human PDLCs, to explore the possibility of using SMFs as a therapeutic agent in periodontal or bone regeneration.
The present study showed that SMF exposure did not affect or impair cell proliferation, even over the course of 14 days of culture. Therefore, the present results agreed with previous findings that demonstrated that SMF exposure did not affect cell proliferation in rat calvaria cells or osteoblast-like cells [12]. However, SMFs significantly inhibited the proliferation of human osteoblasts [13], but conversely stimulated the proliferation of MSCs in vitro [17]. These discrepancies may stem from differences in culture conditions, cell type, and the application methods or type of magnetic fields.
ALP activity is an early maker of osteoblasts, whereas mineralization of the extracellular matrix detected by Alizarin red staining is a marker of mature osteoblasts [26]. OCN and OPN are non-collagenous proteins found in the extracellular matrix produced by osteoblasts, and are late osteoblast differentiation markers [27]. Runx2 is an osteoblast-specific-transcription factor essential for early osteoblastic differentiation [28]. hFOB 1.19 cells have the matrix-synthesizing properties of differentiated osteoblasts, and are an excellent model for the study of osteoblast biology in vitro [29]. We reported above that SMFs increased ALP activity, upregulated OCN, OPN, and Runx2 mRNA, and promoted the formation of mineralized nodules, as well as increasing calcium content in osteoblasts. Evidently, SMFs stimulated both early and late osteoblastic differentiation, as well as mineralization in the osteoblasts, which is consistent with previous data obtained using rat osteoblasts [12], human osteoblast-like MG63 cells [15], dental pulp cells [16], and MSCs [17].
Cementoblasts share many characteristics with osteoblast-like cells, including the ability to form a mineralized matrix, and express genes associated with osteogenic differentiation, including OPN, OCN, CEMP-1, and CAP [22]. PDLCs have the capacity to differentiate in vitro into osteoblasts and cementoblasts, as they have high ALP activity and produce considerable OPN, bone sialoprotein, and OCN [30]. Thus, the regulation of osteogenic/cementoblastic differentiation in PDLCs and cementoblasts has important implications for the development of new therapeutic strategies for periodontitis. We observed that SMFs stimulated PDLCs and cementoblasts to acquire cementoblast/osteoblast characteristics as measured by increased ALP activity, osteogenesis/cementogenesis-related gene expression, enhanced calcium content, and mineral nodule formation. These results support existing data suggesting that SMFs enhance osteoblastic/cementoblastic differentiation and the mineralization of osteoblasts, PDLCs, and cementoblasts.
Wnt/β-catenin signaling plays an important role in skeletal development and osteoblast differentiation [31]. Binding of Wnt to its receptors leads to the inhibition of GSK-3β and the stabilization and accumulation of β-catenin, which then translocates to the nucleus where it interacts with members of the T-cell factor/lymphoid enhancer factor family of transcription factors, resulting in the activation of Wnt downstream target genes [32]. A recent study showed that PEMFs can prevent ovariectomy-induced bone loss and the deterioration of bone microarchitecture and strength by activating this Wnt/β-catenin signaling pathway [33]. In the present study, SMFs significantly upregulated canonical Wnt ligands (Wnt1 and Wnt3a), and stimulated phosphorylation of GSK-3β and total β-catenin expression in osteoblasts. These results suggest that SMFs stimulate osteoblastic differentiation by activating the Wnt/β-catenin signaling pathway.
The MAPK and NF-κB pathways are known to play essential roles in cell differentiation [22]. Recently it was reported that extremely low-frequency magnetic fields inhibited the adipogenic differentiation of human MSCs via the JNK-dependent Wnt signaling pathway [34]. However, electromagnetic fields activated the ERK/MAPK pathway in fibroblast-like cells [35] and hippocampal neurons [36], as well as p38 signaling in a Chinese hamster lung cell line [37]. In addition, treatment with SB203580, an inhibitor of p38 MAPK, suppressed extremely low-frequency electromagnetic field-induced collagen production in mouse osteoblast-like MC3T3-E1 cells [38]. PEMF exposure significantly increased the anti-inflammatory effect of the A2A and A3 adenosine receptors by inhibiting NF-κB pathways in chondrocytes and osteoblasts [39]. In contrast, strong SMFs (6 T) induced the secretion of prostaglandin E2 and the expression of cyclooxygenase 2 in MC3T3-E1 cells, via the translocation of NF-κB [40]. Our results confirm that the p38, JNK/MAPK, and NF-κB signaling pathways are activated in SMF-treated osteoblasts. Furthermore, inhibitors of Wnt, p38, JNK MAPK, and NF-κB significantly reduced osteoblastic differentiation in response to SMFs. Hence, SMF may promote osteoblast differentiation by activating the Wnt, p38, JNK/MAPK, and NF-κB signaling pathways.
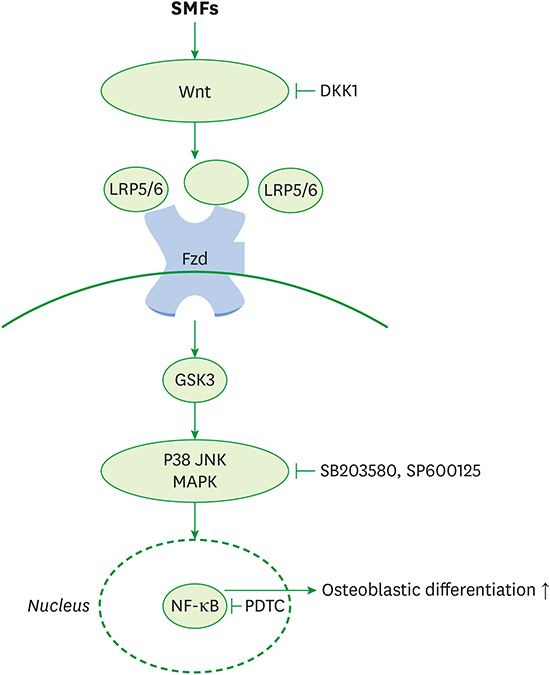
In summary, this is the first study to report that SMFs promoted osteoblastic/cementoblastic differentiation by activating the Wnt/β-catenin, p38, JNK/ MAPK, and NF-κB signaling pathways in osteoblasts, cementoblasts, and PDLCs. Figure 8 is a schematic representation of the signaling pathway involved in the effects of SMFs on the osteoblastic differentiation in osteoblasts. Wnt binds to the transmembrane receptor Frizzled (Fzd) and co-receptor LRP5/6. SMF-activated Wnt-Fzd-LRP5/6 binding events lead to the intracellular accumulation of β-catenin and phosphorylation of GSK3, which activates MAPK and NF-κB for osteoblastogenic gene and Runx2 transcription. Our results also show that SMFs have beneficial effects on bone healing and periodontal regeneration.
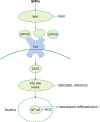 | Figure 8Schematic diagram illustrating the Wnt, Akt, MAPK, and NF-κB signaling pathways triggered by exposure to SMFs, which ultimately stimulate the osteoblastic differentiation of human osteoblasts.
MAPK: mitogen-activated protein kinase, NF-κB: nuclear factor-κB, SMF: static magnetic field, Fzd: Frizzled, GSK-3β: glycogen synthase kinase-3β, JNK: c-Jun N-terminal kinase.
|
 XML Download
XML Download