

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Recent developments in the field of medicine and the consequent increase in the average life expectancy of humans have triggered attempts to enhance and maintain quality of life. Oral and dental health is crucial for consuming a balanced nutritional intake, and is therefore one of the necessities for maintaining quality of life. However, individuals who have lost their teeth for various reasons often need dental treatment, and implants have been used effectively for treating such patients. Consequently, the importance of successful alveolar bone regeneration — one of the requirements for successful implant treatment — has been highlighted.
According to a report by Schropp et al. [1], the width of the buccal, lingual, and palatal alveolar bone decreases by approximately 30% in the initial 3 months after tooth extraction, and approximately 50% of the original width is absorbed 12 months after tooth extraction [2]. Moreover, the bone remaining after tooth loss from periodontal disease is often of poor quality, and the quantity of alveolar bone is not sufficient for implantation. In such cases, alveolar bone regeneration is required prior to implantation to ensure the presence of sufficient alveolar bone for the procedure to be successful. Guided bone regeneration is a commonly used method of alveolar bone regeneration [34].
The 3 key factors required for successful bone regeneration are the presence of osteogenic cells for bone generation, a scaffold for bone conduction, and growth factors for the stimulation of bone regeneration [567]. Growth factors that enhance the efficiency of bone regeneration are currently being actively studied. Growth factors provide a suitable environment for bone regeneration and encourage osteogenic cells to develop near the graft via a cell signaling cascade, eventually promoting alveolar bone regeneration [8].
In this study, we attempted to determine the effects of 1,25-dihydroxyvitamin D3 on bone generation. 1,25-dihydroxyvitamin D3, an active form of vitamin D, is involved in the maintenance of calcium and phosphate homeostasis within the body. In addition, it contributes to various bodily functions, including the maintenance of bone growth, as well as immune and antitumoral functions [9]. The initial step in 1,25-dihydroxyvitamin D3 activation is the conversion of 7-dehydrocholesterol synthesized on the skin to cholecalciferol (vitamin D3) after ultraviolet (UV) exposure; cholecalciferol is then converted into 25-hydroxycholecalciferol by microsomal enzymes in the liver. Hydroxylation of this product by mitochondrial enzymes from the kidney results in the production of 1,25-dihydroxycholecalciferol [1011]. Thus, 1,25-dihydroxycholecalciferol, the final active form, is involved in the maintenance of calcium and phosphate homeostasis within the human body, along with parathyroid hormone and calcitonin. Vitamin D, a fat-soluble vitamin, is considered to be a hormone, as it is biosynthesized from cholesterol and controls gene expression by binding to nuclear vitamin D receptors (VDRs). VDRs are expressed in multiple target cells, including the cells of the small intestine, parathyroid, kidney tubule, and skin epithelium. More specifically, these receptors are expressed in osteoblasts and promote the synthesis of specific proteins, such as osteocalcin (OCN), osteopontin, and collagen type I (Col-I) [1213]. However, whether vitamin D acts directly on osteoblasts and promotes calcification of the bone tissue remains unclear, because the environmental conditions of osteogenesis at the cellular level and at the level of the human body are significantly different. Therefore, we performed an in vitro experiment using MC3T3-E1 osteoblast-like cells derived from mouse calvaria to assess the role of 1,25-dihydroxyvitamin D3 in osteogenesis at the cellular level.
Go to : 
MATERIALS AND METHODS
The cell line used in this study was MC3T3-E1 (an osteoblast-like cell line derived from mouse calvaria) purchased from the American Type Culture Collection (Manassas, VA, USA). The 1,25-dihydroxyvitamin D3 (Bonky®) used in this experiment was purchased from YuYu Pharma, Inc. (Seoul, Korea).
Additionally, α-minimum essential medium (α-MEM) for tissue culture, dimethylsulfoxide (DMSO) for freezing, and trypsin-ethylenediaminetetraacetic acid (EDTA) and antibiotics for cell culture were purchased from Thermo Fisher Scientific (Waltham, MA, USA). The 3-(4,5-dimethylthiazol-2-yl) 2,5-diphenyltetrazolium bromide (MTT) solution for cell proliferation assays, as well as paraformaldehyde and alizarin red S solutions for staining, were purchased from Sigma-Aldrich (St. Louis, MO, USA). The alkaline phosphatase (ALP) assay was performed using the ALP kit (Wako Pure Chemical Industries, Osaka, Japan).
Primers for polymerase chain reaction (PCR) were purchased from Bioneer (Daejeon, Korea). Trizol solution (Molecular Research Center, Cincinnati, OH, USA), chloroform (JUNSEI, Tokyo, Japan), and 2-propanol (Sigma-Aldrich) were used for RNA extraction. Rnasin® Plus RNAse inhibitor and deoxyribonucleoside triphosphates (dNTP) were purchased from Promega (Madison, WI, USA). cDNA was synthesized using a M-MLV cDNA synthesis kit (Enzynomics, Daejeon, Korea), and the DNA ladder was purchased from Nanohelix (Daejeon, Korea).
Cell culture and induction of cell differentiation
MC3T3-E1 cells were cultured in α-MEM (Gibco, Grand Island, NY, USA) with 10% heat-inactivated fetal bovine serum (FBS) and 1% antibiotics (100 U/mL penicillin, 100 µg/mL streptomycin) at 37°C in the presence of ≤5% CO2. Cells were detached using a 0.02% EDTA-0.05% trypsin solution and subcultured when they were 70% confluent. The cell culture media for the negative and positive controls and experimental groups were as follows: α-MEM with 10% FBS for the negative control group; α-MEM with 10% FBS, 10 mM β-glycerophosphate, and 50 µg/mL ascorbic acid to induce differentiation in the positive control group; and the same medium with a serial dilution of 1,25-dihydroxyvitamin D3 in the experimental group. All cells were cultured in the incubator in the presence of 5% CO2 and 95% humidity.
Assessment of cell viability and proliferation using the MTT assay
The MTT assay was used to assess the proliferation of osteoblasts treated with different concentrations (10−4, 10−6, 10−8, 10−10, 10−12, and 10−14 M) of 1,25-dihydroxyvitamin D3 [1415]. Formazan synthesized within the cells was suspended in DMSO, and the proliferation rate was compared by assessing its relative optical density (OD). First, we plated 1×104 cells/well on a 96-well plate and allowed them to grow for 3 days. Different concentrations of 1,25-dihydroxyvitamin D3 were added to the cells when they were 70% confluent, followed by cell culture. Cells in the control group were cultured only in the growth medium, while the other conditions remained constant. On days 1 and 3 of cell culture, 20 µL of 5 mg/mL MTT solution was added to each well, and the medium was removed after 4 hours. Next, 200 µL of DMSO was added to dissolve the synthesized insoluble formazan crystals. The samples were incubated at room temperature for 15 minutes, and OD at 540 nm was measured using a microplate reader (Biotek Instruments, Seoul, Korea). This value of OD represented the amount of MTT reduction by the cells, and was proportional to the number of viable cells in each well. The cell proliferation rates were represented as a percentage value (OD of MTT solution/OD of the control group).
Assessment of ALP activity
MC3T3-E1 cells were plated on a 12-well plate at a density of 2×104 cells/well and cultured until they reached 70% confluency. Cells in the negative control group were cultured in α-MEM growth medium with 10% FBS, whereas the cells in the positive control group were cultured in the same medium with 10 mM β-glycerophosphate and 50 µg/mL ascorbic acid to induce differentiation. The cells in the experimental group were cultured in the same differentiation medium as the positive group, but with different concentrations (10−4, 10−6, 10−8, 10−10, 10−12, and 10−14 M) of 1,25-dihydroxyvitamin D3 [1415]. Cell culture medium was collected on days 7 and 14, and ALP activity was measured using an ALP kit (Wako Pure Chemical Industries). The measurements were represented as a percentage value in comparison to the measurements from the positive group after excluding the blank values.
Separation and quantification of total RNA
The MC3T3-E1 cells were divided into a positive control group and an experimental group. Cells were grown in the growth medium with 10−12 M 1,25-dihydroxyvitamin D3, which exhibited the highest level of ALP activity, for 7 and 14 days, and the RNA was extracted. We assessed the expression of Col-I, OCN, and ALP, which were expressed during osteoblast differentiation, as well as those of VDRs, to understand the molecular mechanism of 1,25-dihydroxyvitamin D3 using reverse transcription (RT)-PCR. Cultured cells were washed with diethyl pyrocarbonate (DEPC) water, and RNA was extracted using 1 mL of TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Then, 200 µL of chloroform was added to the isolated RNA solution and the reaction mixture was incubated on ice. Next, the mixture was centrifuged at 12,000 rpm, and 500 µL of supernatant was separated. RNA was precipitated by the addition of 0.5 mL of isopropyl alcohol and incubation at −20°C for 12 hours, followed by centrifugation at 12,000 rpm for 10 minutes. The supernatant was removed, and the pellets were washed with 80% ethanol and air-dried. RNA was dissolved in 30 µL of RNase-free water, and the solution was heated to 60°C prior to the measurement of OD260. The extracted RNA was quantified and stored at −80°C.
We added the same amount of 2-propanol as the supernatant, mixed the solution, and then re-centrifuged at 12,000 rpm for 10 minutes. The supernatant was removed, and the pellets were washed using 75% ethanol and air-dried. RNA was dissolved in 30 µL RNase-free water and quantified after heating at 60°C, followed by storage at −80°C.
RT-PCR
An RT-PCR kit (Invitrogen) was used for the RT-PCR analysis. We added 2 µL of dNTPs, 2 µL of dithiothreitol (DTT), 4 µL of reaction buffer, 1 unit/µL reverse transcriptase, 0.5 µL of RNase inhibitor, and 8.5 µL of DEPC to 1 µL (1 µg RNA) of total RNA solution extracted from the control and experimental groups. The solution was mixed and incubated for 1 minutes at 37°C. First, the samples were incubated at 4°C for approximately 5 minutes to allow random primer annealing. Next, the RT buffer, dNTP, RNase inhibitor, DTT (100 nM), and reverse transcriptase (M-MLV, 200 units/µg) were added to the solution and RT was performed at 42°C for 90 minutes. The reaction mixture was incubated at 95°C for 5 minutes, and the synthesized cDNA was used as a template for quantitative PCR (qPCR). PCR was performed using a master mix of 10X PCR buffer, 10 mM dNTPs, 10 pmol of glyceraldehyde 3-phosphate dehydrogenase (GAPDH), ALP, OCN, Col-I, and vitamin D receptor (VDR) primers, the cDNA, and 1 unit of Taq polymerase (Promega) in a total reaction volume of 20 µL. The primer sequences and reaction conditions were optimized for each gene (Table 1). The PCR conditions were as follows: 3 minutes of pre-denaturation at 94°C, 30 seconds of denaturation at 94°C, 42 seconds of annealing at 58°C, 30 seconds of extension at 72°C (35 cycles), and 10 minutes of final extension at 72°C. The Tm of each primer was considered before finalizing the annealing temperature at 55°C for all primers. The amplified PCR products were electrophoresed on 1.2% agarose gel after mixing the product with 6× DNA loading dye. The gel was stained with ethidium bromide and imaged under UV. The band densities were assessed using the Image J program (National Institutes of Health, Bethesda, MD, USA) and represented graphically after normalizing to GAPDH levels.
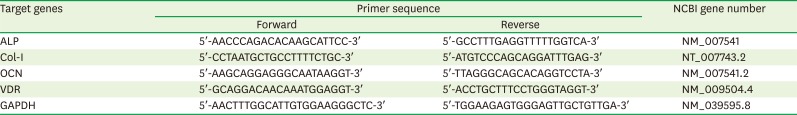
Table 1
Primer sequences for RT-PCR
RT-PCR: reverse transcription-polymerase chain reaction, NCBI: National Center for Biotechnology Information, ALP: alkaline phosphatase, Col-I: collagen type I, OCN: osteocalcin, VDR: vitamin D receptor, GAPDH: glyceraldehyde 3-phosphate dehydrogenase.
![]()
Alizarin red S staining
Alizarin red S staining was performed to assess the level of bone differentiation. MC3T3-E1 cells were plated on a 12-well plate at a density of 2×104 cells/well, and cultured to 70% confluency. Cells in the negative control group were cultured in α-MEM growth medium with 10% FBS, whereas the cells in the positive control group were cultured in the same medium with 10 mM β-glycerophosphate and 50 µg/mL ascorbic acid to induce differentiation. Cells from the experimental group were cultured in growth media containing 10−12 M 1,25-dihydroxyvitamin D3, which exhibited the highest level of ALP activity. The cells were fed with new media every 3 days and staining was performed after 3 weeks. The medium was removed, and the cells were washed thrice with 1 mL of Dulbecco's phosphate-buffered saline (DPBS) for 5 minutes per wash. Next, the cells were fixed with 4% paraformaldehyde for 20 minutes at room temperature, following which the fixation solution was removed by washing with distilled water (≥2×, 10 minutes per wash). The cells were stained using 2% alizarin red S (pH 4.2) for 10 minutes at room temperature, with minimal light exposure. The stained cells were washed again with DPBS, and assessed using a light microscope.
Statistical analysis
The results from this study were represented as mean±standard deviation, with minimum of 5 replicates. Statistical analysis was performed using SPSS version 20.0 (IBM Corp., Armonk, NY, USA). The Mann-Whitney test was used to assess the differences between the positive control group and experimental groups at a significance level of 0.05 with 95% confidence intervals.
Go to :
RESULTS
Assessment of cell viability
We performed MTT assays to elucidate the effects of 1,25-dihydroxyvitamin D3 on MC3T3-E1 proliferation. Compared to the cell viability of the positive group (100.00±0.25), there were no significant differences on day 1, despite the slightly lower viability observed for cells treated with most of the 1,25-dihydroxyvitamin D3 concentrations (except for 10−4, 10−8, and 10−12 M). However, compared to the cell viability of the positive group (100%±0.031%) on day 3, the viability of the experimental group increased by 9%–12%, and it was significantly higher for cells treated with 10−4, 10−8, and 10−12 M 1,25-dihydroxyvitamin D3 (n=6, P<0.05) (Figure 1). Thus, based on the results obtained on day 3, we conclude that 1,25-dihydroxyvitamin D3 positively affected cell proliferation.
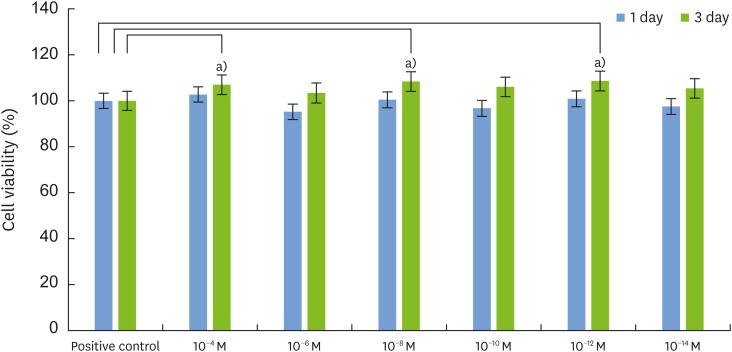 | Figure 1Effects of 1,25-dihydroxyvitamin D3 on the proliferation of MC3T3-E1 cells. The cells were cultured in the presence of various concentrations of 1,25-dihydroxyvitamin D3 for 1 and 3 days. Cell viability was measured by the MTT assay. Cells cultured in growth medium alone served as the control (n=6). The results are expressed as the percentage of the control values, and the data are the mean±SD values from 3 independent experiments.
MTT: 3-(4,5-dimethylthiazol-2-yl) 2,5-diphenyltetrazolium bromide, SD: standard deviation.
a)Statistically significant difference (n=6, P<0.05 as compared with the control).
|
Assessment of ALP activity
We analyzed ALP activity to assess the level of osteoblast differentiation. We assessed ALP activity on days 7 and 14 after adding different concentrations of 1,25-dihydroxyvitamin D3 (10−4, 10−6, 10−8, 10−10, 10−12, and 10−14 M) to MC3T3-E1 cells. Compared to the positive control group, ALP activity increased in the experimental groups treated with 10−8, 10−10, 10−12, and 10−14 M 1,25-dihydroxyvitamin D3 on day 7. More specifically, ALP activity significantly increased at low concentrations (10−10, 10−12, and 10−14 M) (n=6, P<0.05). Compared to the positive control group, ALP activity was significantly higher in the experimental groups treated with 10−10, 10−12, and 10−14 M 1,25-dihydroxyvitamin D3 on day 14 (n=6, P<0.05). ALP activity was highest in cells treated with 10−12 M 1,25-dihydroxyvitamin D3 (Figure 2).
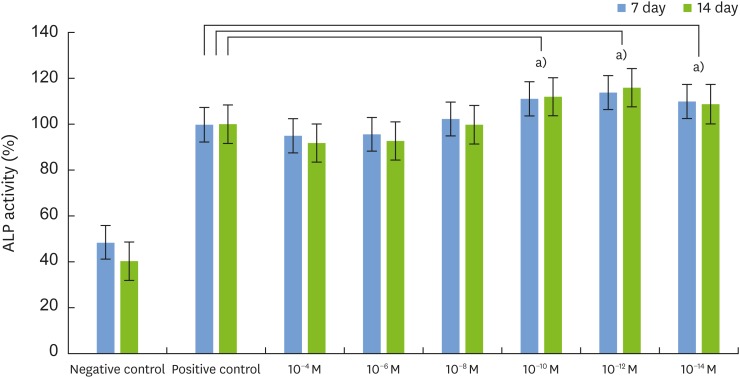 | Figure 2Effects of 1,25-dihydroxyvitamin D3 on ALP activity in MC3T3-E1 cells. Cells were cultured in differentiation medium (positive control) or in the presence of 10−4, 10−6,10−8,10−10,10−12, and 10−14 M 1,25-dihydroxyvitamin D3, respectively. Cell viability was measured by the MTT assay. Cells cultured in growth medium alone served as the negative control. Results were expressed as the percentage of the control values, and the data are the mean±SD values from 3 independent experiments.
ALP: alkaline phosphatase, MTT: 3-(4,5-dimethylthiazol-2-yl) 2,5-diphenyltetrazolium bromide, SD: standard deviation.
a)Statistically significant difference (n=6, P<0.05 compared to the positive control).
|
Osteogenesis-related gene expression analysis
Expression of Col-I mRNA
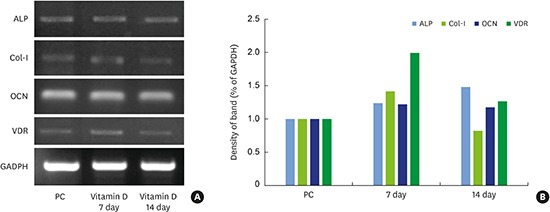
We assessed the levels of Col-I mRNA after 1,25-dihydroxyvitamin D3 treatment using RT-PCR. The cells used in this experiment were treated with 10−12 M 1,25-dihydroxyvitamin D3, which exhibited the highest level of cell proliferation and differentiation based on the MTT and ALP assays. Compared to the positive control group (100%), the experimental group showed higher Col-I mRNA levels (142%) on day 7 post-differentiation, whereas its expression decreased to 83% on day 14 post-differentiation (Figure 3).
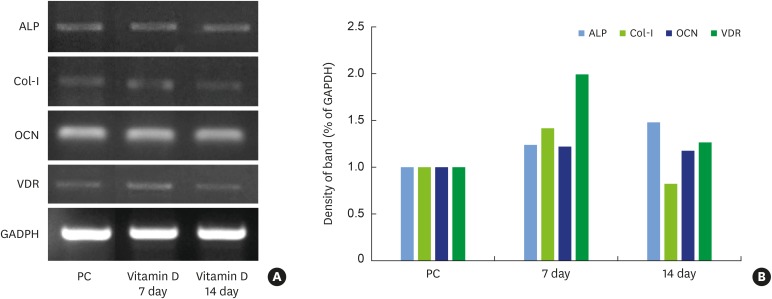 | Figure 3RT-PCR analysis of osteogenesis-related mRNA expression in MC3T3-E1 cells treated with 1,25-dihydroxyvitamin D3 (A). GAPDH was used as the internal control. The percentage of osteogenesis-related gene expression was calculated relative to that of GAPDH (B).
RT-PCR: reverse transcription-polymerase chain reaction, GAPDH: glyceraldehyde 3-phosphate dehydrogenase, PC: positive control, ALP: alkaline phosphatase, Col-I: collagen type I, OCN: osteocalcin, VDR: vitamin D receptor.
|
Expression of ALP mRNA
Compared with the positive control group (100%), ALP mRNA expression levels increased to 124% in the experimental group on day 7 post-differentiation, and further increased to 148% on day 14. Thus, ALP mRNA expression continued to increase until day 14 post-differentiation after 1,25-dihydroxyvitamin D3 treatment (Figure 3).
Expression of OCN mRNA
OCN mRNA expression slightly increased in the 1,25-dihydroxyvitamin D3-treated group (122%) on day 7 post-differentiation. Although a slight decrease from the day 7 level was observed on day 14 (118%), this difference was not statistically significant. Thus, OCN mRNA levels in the positive control group and experimental groups were similar, and the change in expression levels through day 14 post-differentiation was negligible (Figure 3).
Expression of VDR mRNA
VDR mRNA levels increased 2-fold (199%) in the 1,25-dihydroxyvitamin D3-treated group compared to the positive control group (100%) on day 7 post-differentiation. However, the mRNA level then decreased (127%) on day 14 compared to the level on day 7. Therefore, we conclude that VDR expression dramatically increased through day 7 (the initial phase of differentiation), and started to decrease after the middle phase of differentiation (Figure 3).
Analysis of calcified nodules
We performed alizarin red S staining to assess the level of calcified nodule formation in MC3T3-E1 cells and further observed the nodules under a light microscope. Observation with the naked eye showed minimal staining in the negative control group on days 7, 14, and 21, but increased staining was observed in both the positive control and experimental groups treated with 10−12 M 1,25-dihydroxyvitamin D3. The visual differences between the positive control group and the experimental groups were not significant, and both groups exhibited increased levels of staining over time (Figure 4A).
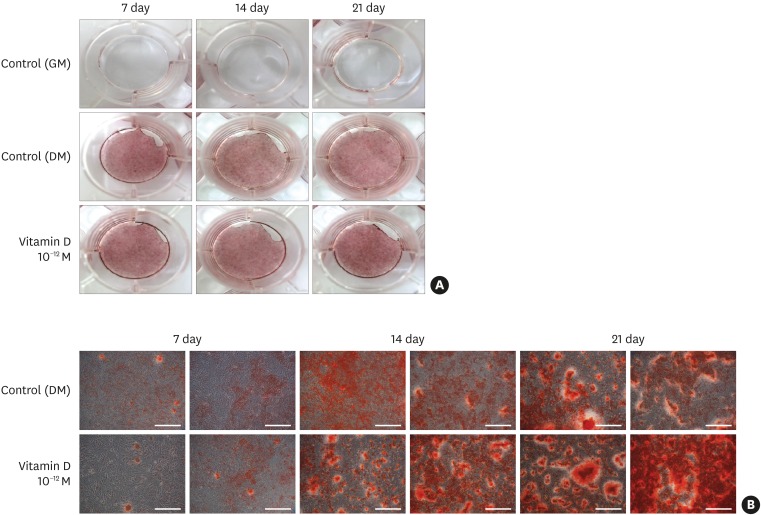 | Figure 4Effects of 1,25-dihydroxyvitamin D3 on the formation of mineralized nodules in MC3T3-E1 cells. Cells were observed as macroscopic images (A). Effects of 1,25-dihydroxyvitamin D3 on the formation of mineralized nodules in MC3T3-E1 cells. (B) Cells were cultured for 7, 14, and 21 days in DM or in the presence of 10−12 M 1,25 dihydroxyvitamin D3 (DM + 1,25-vitamin D). The negative control cells were cultured in GM alone (scale bar=250 μm).
DM: differentiation medium, GM: growth medium.
|
When observed at ×40 under a light microscope, the formation of calcified nodules was more pronounced in the 1,25-dihydroxyvitamin D3-treated group than in the positive control group in the initial phase of differentiation. On days 7, 14, and 21, both groups exhibited an increased number and size of calcified nodules, but the difference was not evident with the naked eye. However, the experimental group tended to develop calcified nodules earlier than the positive control group (Figure 4B).
Go to :
DISCUSSION
Interest in implants as a treatment for restoring lost teeth is increasing with the worldwide rise in life expectancy. However, the absorption of alveolar bone due to chronic periodontal diseases or shrunken alveolar bone resulting from long-term use of dentures renders implantation a challenging prospect. In these cases, pre-implantation treatments are required to restore the lost alveolar bone, and among the many treatments that have been developed, guided bone regeneration has been frequently used to restore the alveolar ridge [16].
Osteogenic cells (for bone generation), a scaffold (for bone conduction), and growth factors (for stimulation of bone regeneration) are required to regenerate the alveolar bone [17]. An adequate cell model is required to study the development and differentiation of osteoblasts. The MC3T3-E1 osteoblast-like cells used in this study are osteoprogenitor cells derived from mouse calvaria that are capable of bone generation, and are therefore widely used in bone biology. These cells have similar metabolic characteristics to osteoblasts in terms of proliferation, differentiation, and calcification. Therefore, they are ubiquitously used in studies that assess the differentiation of osteoblasts [18].
Osteogenesis largely consists of the proliferation and differentiation of osteoblasts, the formation of extracellular matrix, and mineralization [19]. Factors associated with the cell cycle, such as histones and ribosomal proteins, are present at high concentrations during proliferation and differentiation. Extracellular matrix proteins occur in 2 different patterns at the extracellular matrix formation and maturation stages. First, Col-I, ALP, and transforming growth factor beta 1 occur in abundance during the maturation stage. During the mineralization stage, proteins such as fibronectin, osteonectin, and OCN are highly expressed. The activation of ALP, which is often used as a biomarker of bone differentiation, occurs at high levels in the early stages of extracellular matrix formation, but decreases with time. Bone nodules are formed and OCN is expressed at high levels during the mineralization period of the bone matrix [20].
During osteoblast development, growth factors provide an adequate environment for bone regeneration and stimulate osteoblasts via cellular signaling cascades. Several studies on growth factors, including bone morphogenetic protein 2, platelet-derived growth factor, transforming growth factor beta, insulin-like growth factor, and glucocorticoids, have confirmed that growth factors positively affect osteogenesis [21]. The goal of this study was to assess the effects of 1,25-dihydroxyvitamin D3, which is involved in maintaining calcium and phosphate homeostasis in the human body during osteogenesis, on osteoblasts. The metabolite 1,25-dihydroxyvitamin D3 is a fat-soluble vitamin similar to vitamins A, E, and K, and it has been identified as important in several studies on metabolism. 1,25-dihydroxyvitamin D3 is biosynthesized from cholesterol by the cytochrome P450 enzyme. It binds to VDRs within the nucleus, and controls nuclear transcription. This mechanism of action is similar to those of steroids, and therefore 1,25-dihydroxyvitamin D3 is classified as a hormone [12]. VDRs are expressed in several target cells, including those of the small intestine, parathyroid, kidney tubule, and skin epithelium. Specifically, in the bone tissue, VDRs are expressed in osteoblasts. In osteoblasts or osteoblast-like cells that express VDRs, 1,25-dihydroxyvitamin D3 is known to promote the expression of specific proteins, such as collagen, OCN, osteopontin, and matrix Gla [2223242526]. Briefly, the mechanism of 1,25-dihydroxyvitamin D3 includes the formation of a complex between 1,25-dihydroxyvitamin D3 and a VDR, followed by recognition of the vitamin D responsive elements in the DNA to activate the transcription of several related genes [27].
However, whether vitamin D directly affects osteoblasts in localized environments to promote differentiation and mineralization remains unclear. Therefore, we assessed whether 1,25-dihydroxyvitamin D3 affected bone tissue formation and mineralization at the osteoblast level in vitro.
We first performed the MTT assay to understand the effects of vitamin D on the proliferation of osteoblasts. A comparison of cell proliferation on days 1 and 3 between the positive control group and the experimental group (treated with 1,25-dihydroxyvitamin D3) demonstrated no negative effects of 1,25-dihydroxyvitamin D3 on cell proliferation. On day 3, some of the experimental groups exhibited slightly higher cell proliferation than the positive control group. All experimental groups exhibited higher cell viability than the positive control group (100%±0.031%), and the levels were significantly higher for 10−4, 10−8, and 10−12 M concentrations (P<0.05). Therefore, we concluded that 1,25-dihydroxyvitamin D3 did not negatively affect the proliferation of MC3T3-E1 osteoblast-like cells; instead, it positively affected cell proliferation on day 3.
We next measured ALP activity to assess the level of differentiation of the osteoblast-like cells. Compared to the positive control group (P<0.05), ALP activity increased in cells treated with relatively low concentrations (10−10, 10−12, and 10−14 M) of 1,25-dihydroxyvitamin D3, and the activity level was significantly higher at these concentrations on days 7 and 14 post-treatment. The differentiation of MC3T3-E1 cells can be divided into 2 major stages. The first stage (10–15 days after differentiation) involves the expression of specific proteins, such as ALP, and the maturation of the extracellular matrix. The second stage (25–30 days after differentiation) involves the expression of proteins such as OCN and the mineralization of the extracellular matrix [28]. In this study, we demonstrated that on days 7 and 14, ALP activity continued to increase, indicating that these osteoblast-like cells underwent normal differentiation. ALP levels were lower in the experimental groups treated with higher concentrations of 1,25-dihydroxyvitamin D3. A previous study by Jones [29] in 2008 reported that high concentrations of 1,25-dihydroxyvitamin D3 induced the production of analogous compounds, such as 24,25(OH)2D3, 25,26(OH)2D3, and 25(OH)D3-26,23-lactone, which compete with 1,25-dihydroxyvitamin D3 to prevent the binding of vitamin D to its receptors. Therefore, to increase the efficiency of differentiation in osteoblasts, an adequate concentration of 1,25-dihydroxyvitamin D3 must be maintained at the correct locations for the appropriate amount of time. Consequently, additional studies on the development of effective carriers that can function as described above should be performed in the future [30].
We performed RT-PCR to assess the changes in the expression of several candidate genes after 1,25-dihydroxyvitamin D3 treatment. The mRNA levels of ALP, Col-I, VDRs, and OCN were higher in the experimental group than in the positive control group on day 7, which represented the early phase of differentiation. ALP expression slightly increased on day 14, but Col-I and VDR expression decreased. In 1986, Kurihara et al. [31] showed that 1,25-dihydroxyvitamin D3 had a stimulatory effect during the early phase of differentiation in MC3T3-E1 cells. More specifically, they reported that Col-I and ALP levels increased, and that VDRs played a role in the elevation of these protein levels. The authors also emphasized that VDR expression was greatly upregulated on day 7 after 1,25-dihydroxyvitamin D3 treatment. Thus, 1,25-dihydroxyvitamin D3 may be involved in upregulating VDR expression in osteoblasts, and 1,25-dihydroxyvitamin D3 and VDR form a complex to activate the transcription of several genes within the nucleus. Kurihara et al. [31] also reported that during the initial phase of differentiation, active synthesis of Col-I resulted in the formation of the extracellular matrix, and that ALP was deposited in the extracellular matrix to assist in the maturation process. The results from our study were similar, as Col-I expression increased on day 7 (initial stage) in the experimental group, followed by a decrease on day 14, whereas ALP expression continued to increase until day 14. OCN expression increased on both days 7 and 14 in the experimental group treated with 1,25-dihydroxyvitamin D3; however, the level was not significantly different from that of the positive control group. During the differentiation of MC3T3-E1 cells, the expression of proteins such as OCN dramatically increased during the later period (18–23 days after differentiation), followed by matrix mineralization. Considering that days 7 and 14 correspond to the early stages of cellular differentiation and matrix maturation, it is natural that OCN expression did not increase dramatically [29].
We performed alizarin red S staining to understand the effects of 1,25-dihydroxyvitamin D3 on the last step of osteogenesis, namely, matrix mineralization. Calcium and phosphate in the extracellular matrix are crucial for the formation and mineralization of bone nodules, which are bright red in color after alizarin red S staining. The levels of deposited calcium and phosphate increased with time, and the stained bone matrix was visible at 14 days after differentiation [32]. Similarly, in this study, we began to observe the stained bone matrix at 14 days after differentiation, and it continued to increase in quantity until day 21. Overall, the level of mineralization was not different between the positive control and the experimental groups; however, bone nodules were observed earlier in the experimental group than in the positive control group. These results are in agreement with those of a previous study by Wang et al. [33], wherein the authors showed that the injection of growth factors resulted in earlier formation of bone nodules. Therefore, if 1,25-dihydroxyvitamin D3 is injected along with the bone graft material to promote earlier bone nodule formation during local guided bone regeneration, the early stages of osteogenesis, which are normally unstable, may be promoted and the efficiency of the general osteogenesis process may also be improved.
In this study, we assessed the effects of 1,25-dihydroxyvitamin D3 on the proliferation, differentiation, and mineralization of osteoblasts at the cellular level. 1,25-dihydroxyvitamin D3 exhibited no inhibitory effects on cell proliferation at any of the tested concentrations and showed positive effects at optimal concentrations. Moreover, treatment with optimal concentrations of 1,25-dihydroxyvitamin D3 resulted in higher ALP activity, and the mRNA levels of different proteins (Col-I, ALP, VDRs, and OCN) increased compared to the positive control group. The alizarin red S assay showed that treatment with 1,25-dihydroxyvitamin D3 had no significant effect on bone mineralization, but induced early bone nodule formation. In conclusion, our results demonstrated that 1,25-dihydroxyvitamin D3 can promote proliferation, differentiation, and mineralization in MC3T3-E1 cells.
Go to :
 XML Download
XML Download