

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Cell therapy has emerged as an advanced medical technology for restoring damaged tissues and organs. Of the various cell sources, mesenchymal stem cells (MSCs) have been used most frequently in experiments and in clinical trials in the field of regenerative medicine. In addition to the effects of the cell types into which the MSCs could differentiate, the MSCs themselves have been shown to exert paracrine actions that promote the repair of damaged tissues.1234 There is increasing evidence that cell therapy produces positive effects largely via the secretion of various cytokines, such as growth factors, cytokines and chemokines with anti-scarring, anti-apoptotic, angiogenesis, immunomodulatory, and chemoattractant functions.1
There have been suggestions that mononuclear cells (MNCs) might also be used to regenerate tissues because they can be isolated and cultured from the MNC fractions of bone marrow (BM), cord blood (CB), and granulocyte-colony stimulating factor (G-CSF)-mobilized peripheral blood (mPB). The effects of MSCs and MNCs have been compared in various experimental settings.567 Furthermore, in clinical trials of intravenous CB MNCs in children with cerebral palsy (CP) some patients gave partial responses without any significant toxicity.89
However, there have been few reports of the paracrine effects of intravenously infused MNCs in improving neurological functions. This work was aimed to measure the serial cytokine changes in children with CP who participated in a clinical trial of cell therapy with G-CSF followed by autologous mobilized peripheral blood mononuclear cells (mPBMCs).
METHODS
Study subjects
Peripheral blood (PB) samples were collected from 16 CP children who participated in a randomized, double-blind, cross-over clinical study of intravenous infusion of G-CSF followed by mPBMCs. The study design is schematized as follows. Fifty-seven children with the non-severe type of CP were enrolled. After baseline studies (M0), intravenous G-CSF was administered for 5 days, at which time mPBMCs were collected by apheresis and cryopreserved. One month later (M1), the children were randomized for infusion of mPBMCs or placebo. After a further 6 months, cross-over infusion was performed and the children were observed for another 6 months. PB samples for cytokine measurements were collected at baseline (M0), before infusion of mPBMC/placebo (M1, M7) as well as at the end of the study (M13).
Sample preparation and measurement of cytokines
The PB samples were centrifuged at 1,800 rpm for 30 minutes and plasma was separated and stored at −80°C until the end of the clinical study. Thereafter, it was thawed and the following cytokines were measured using a Human Quantikine ELISA kit (R & D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions; G-CSF, brain-derived neurotrophic factor (BDNF), vascular endothelial growth factor (VEGF), insulin-like growth factor (IGF)-1, interleukin (IL)-6, IL-8, IL-10.
Statistical analysis
The cytokine levels were compared between T1 (mPBMC infusion 1 month after G-CSF infusion, n = 8) and T7 (mPBMC infusion 7 months after G-CSF infusion, n = 8) groups, mPBMC and placebo groups, clinical responders and non-responders. Clinical responses were assessed by a rehabilitation medicine specialist according to scores in various evaluation tools such as Gross Motor Function Measure (GMFM), Gross Motor Function Classification System (GMFCS), Manual Ability Classification System (MACS), Denver Developmental Screening Test (DDST), Pediatric Evaluation of Disability Inventory (PEDI). The patients were classified as responders and non-responders according to their clinical outcomes. Clinical responders were defined as having score changes of GMFM > 4 points and score changes in PEDI of > 7 points in 3 items.
Groups were compared using the Wilcoxon rank sum test and Wilcoxon signed rank test, which are nonparametric methods, because there were not many enrolled patients. All statistical analyses were conducted with SAS software (version 9.3; SAS Institute Inc., Cary, NC, USA). P values < 0.05 were accepted as statistically significant.
RESULTS
Changing patterns of cytokine levels in T1 and T7 groups
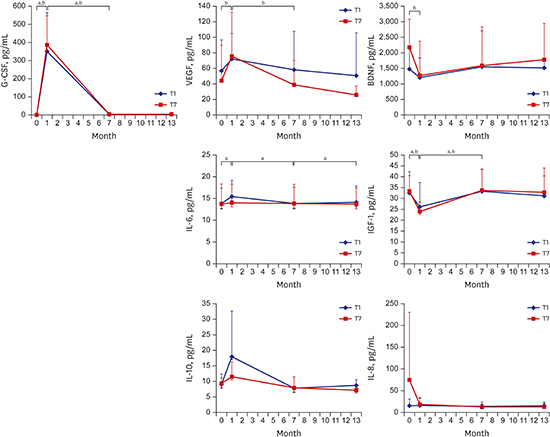
G-CSF levels increased significantly in T1 at 1 month after G-CSF infusion and decreased to baseline levels at 7, 13 months. VEGF, IL-6, and IL-10 levels underwent similar changes and even the VEGF level in the T7 group, like the G-CSF level, was reduced at 7 months, while that of the T1 group was not. On the other hand, BDNF and IGF-1 levels followed the opposite pattern to G-CSF; they decreased significantly at 1 month after G-CSF infusion and recovered at 7 months. Apart from IL-6, which increased after mPBMC infusion at 7 months in the T1 group, there were no changes of cytokine levels following mPBMC infusion at 7 months (Fig. 1).
 | Fig. 1Serum cytokine levels according to the time of infusion of mPBMC in CP children who received intravenous infusion of G-CSF-mPBMCs or placebo. G-CSF levels increased significantly in T1 at 1 month after G-CSF infusion and decreased to baseline levels at 7, 13 months. VEGF, IL-6, and IL-10 levels underwent similar changes and even the VEGF level in the T7 group, like the G-CSF level, was reduced at 7 months, while that of the T1 group was not. On the other hand, BDNF and IGF-1 levels followed the opposite pattern to G-CSF; they decreased significantly at 1 month after G-CSF infusion and recovered at 7 months. Apart from IL-6, which increased after mPBMC infusion at 7 months in the T1 group, there were no changes of cytokine levels following mPBMC infusion at 7 months.
G-CSF = granulocyte-colony stimulating factor, BDNF = brain-derived neurotrophic factor, VEGF = vascular endothelial growth factor, IGF = insulin-like growth factor, IL = interleukin, T1 = PBMC infusion at 1 month after G-CSF infusion, T7 = PBMC infusion at 7 months after G-CSF infusion, CP = cerebral palsy, mPBMC = mobilized peripheral blood mononuclear cell.
P values < 0.05 were accepted as statistically significant in aT1 and bT2 groups.
|
Cytokine levels in clinical responders versus non-responders
When the responders and non-responders were compared, IL-6 (P = 0.050) and G-CSF (P = 0.001) were higher in the responders than the non-responders at 1 month after G-CSF infusion, while BDNF (P = 0.030) and IGF-1 (P = 0.001) were lower. Interestingly, BDNF was higher at baseline in the responders than the non-responders (P = 0.030) (Fig. 2).
 | Fig. 2Serum cytokine levels in the clinical responder/non-responder groups in CP children who received intravenous infusion of G-CSF-mPBMCs or placebo. IL-6 and G-CSF were higher in the responders than the non-responders at 1 month after G-CSF infusion, while BDNF and IGF-1 were lower. Interestingly, BDNF was higher at baseline in the responders than the non-responders.
G-CSF = granulocyte-colony stimulating factor, BDNF = brain-derived neurotrophic factor, VEGF = vascular endothelial growth factor, IGF = insulin-like growth factor, IL = interleukin, CP = cerebral palsy, mPBMC = mobilized peripheral blood mononuclear cell.
P values < 0.05 were accepted as statistically significant in aresponder and bnon-responder groups.
|
Cytokine levels in the mPBMC and placebo groups
To reveal the effect of mPBMCs infusion on the cytokines, the analysis was performed during 6 months after infusion of mPBMCs or placebo. However, there were no significant differences of cytokine levels over 6 months between the mPBMC and placebo groups (Fig. 3).
 | Fig. 3Serum cytokine levels after randomization of CP children who received intravenous infusion of G-CSF-mPBMCs or placebo. There were no significant differences of cytokine levels over 6 months between the mPBMC and placebo groups.
G-CSF = granulocyte-colony stimulating factor, BDNF = brain-derived neurotrophic factor, VEGF = vascular endothelial growth factor, IGF = insulin-like growth factor, IL = interleukin, CP = cerebral palsy, mPBMC = mobilized peripheral blood mononuclear cell.
|
DISCUSSION
Both G-CSF and its receptor are widely expressed by neurons in the central nervous system, and their expression is induced by ischemia, which points to an autocrine protective signaling mechanism. Thus, G-CSF has been revealed to be an endogenous ligand in the CNS that has a dual activity countering acute neuronal degeneration and contributing to long-term plasticity after cerebral ischemia.10
G-CSF also has a neuroprotective role via Th2 switching and regulatory T cell production. There is compelling evidence that it exerts profound immunoregulatory effects by polarizing T cell differentiation via the production of IL-4 and IL-10, which is accompanied by reduced production of interferon-γ and IL-2.11121314 Deboy et al.15 showed that IL-4 is important for facial motor neuron survival after nerve injury, and Frenkel et al.16 also revealed a role of IL-10 produced by CD4+ T cells in neuroprotection. The cytokine IL-6 is also recognized as a trophic factor in the maintenance of normal neurons and in promoting neuronal survival and angiogenesis.17 Ohki et al.18 observed that G-CSF increased plasma VEGF release from neutrophils in vivo together with an increase in circulating neutrophils. The emerging evidence for an etiologic role of VEGF in (at least some types of) neurodegeneration provides the rationale for considering the therapeutic potential of VEGF in neurodegenerative disorders, which are mostly incurable. In this study, the production of VEGF, IL-6, and IL-10 increased in the first month after G-CSF infusion, along with a similar pattern for G-CSF, even after harvesting most of mPBMCs. Based on this study and the previous data regarding the immunoregulatory effects of G-CSF, it appears that G-CSF-derived cytokines as well as G-CSF itself may promote regenerative processes countering neurodevelopmental dysfunction.
BDNF plays a significant role in neurogenesis. It can promote the protective effects of neural stem cells that contribute to the brain's neurogenic response by enhancing their survival.19 IGF-1 also plays a significant role in neuronal development,2021 recovery from neuronal injury,2223 neuronal survival,24 and neurite outgrowth following crush injury.25 In vitro studies have suggested that IGF-1 is produced locally by non-neuronal cells following injury, and that it stimulates regeneration.26 Although BDNF is more concentrated in brain tissue, it is present in the bloodstream and derives from a variety of sources, including platelets and the brain.272829 There have been reports that BDNF can cross the blood-brain barrier30 and a positive correlation between peripheral BDNF protein levels and brain levels have been reported in rodents,31 suggesting that peripheral BDNF levels may reflect BDNF levels in the brain. BDNF blood levels have also been shown to correlate with cortical integrity.32 Hence in clinical settings, PB levels (i.e., serum or plasma) are widely used as a proxy for central levels. However, we observed that G-CSF lowered plasma levels of BDNF and IGF-1, which is not consistent with previous studies that demonstrated that BDNF and IGF-1 were released from endothelial cells activated by G-CSF, and that they contributed to neural regeneration.3334 Therefore, further studies are required to reveal the relation between plasma levels of G-CSF and BDNF/IGF-1.
Other studies have shown that CD34+ cells mobilized by G-CSF can home from the circulating blood to ischemic brain tissues and contribute to the improvement of neurological functions.35 In addition, we recently observed, in a previous study, that G-CSF modified the intracellular expression of inflammatory cytokines in G-CSF-mPBMCs of CP children, and that IL-6 levels were significantly elevated in mPBMCs rather than in PBMCs.36 However, in this study, against our expectations, there were no significant changes of cytokine secretion following booster mPBMC infusion compared to placebo. Nevertheless, VEGF levels did not decrease in the T1 group which received mPBMC at 1 month, but even decreased significantly at 7 months in the same pattern as G-CSF in the T7 group. Therefore, we tentatively suggest that mPBMC may have a booster effect, particularly when infused during elevated G-CSF levels.
We also observed, in the clinically responding group, that IL-6 as well as G-CSF were elevated at 1 month after G-CSF infusion, along with reduced BDNF and IGF-1 levels. In addition, BDNF levels were significantly higher at baseline in the responders than the non-responders. These findings suggest that clinical responses to G-CSF infusion in CP patients could be predicted by measuring BDNF before treatment, and serial changes of IL-6, BDNF, and IGF-1 after treatment.
In summary, the changes of G-CSF itself, as well as G-CSF-induced cytokines such as IL-6, may be associated with the clinical improvement of neurologic functions. The G-CSF-induced changes of IL-6, BDNF, and IGF-1, and BDNF levels before treatment, could be used as prognostic factors in G-CSF trials in CP children. Although a definite effect of mPBMC reinfusion was not detected in this study, further study is needed to clarify the effect of G-CSF or mPBMCs on their own in CP children.
 XML Download
XML Download