

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Asthma is a major health concern in all age groups, and the allergic form of asthma occurs due to a wide range of factors, including exposure to molds, pet dander, cockroaches, and ragweed.12 Early-life exposure to and sensitization with various allergens, including cockroaches and Alternaria, have been noted in children with severe forms of asthma.34 Cockroaches produce several allergens, and exposure to increased levels of these allergens is a major risk factor in sensitized individuals.56 Previously, it was reported that cockroach allergens exhibit proteolytic activity, increasing inflammatory cytokines in human airway epithelial cells, or a Toll-like receptor (TLR) 2 agonist that directly affects neutrophils, inducing an early innate immune response.7 Furthermore, it was shown that cockroach frass proteases can cleave pro-matrix metalloproteinase (MMP)9, which plays a role in airway remodeling.8
The MMP family includes zinc- and calcium-dependent endopeptidases,9 which can degrade a variety of substrates, and they play crucial roles in the regulation of tissue remodeling and regeneration.10 The production and activity of MMPs are tightly controlled by transcriptional and post-translational mechanisms in combination with proenzyme activation and inhibition of active MMPs. However, when not tightly regulated, MMPs can become dangerous and have been shown to be involved in cancer metastasis and cardiovascular diseases. Recent studies revealed that MMP dysregulation occurs in chronic obstructive pulmonary disease, asthma, interstitial lung diseases, and lung cancer.11 MMP levels and/or activity in individuals with asthma were shown to be significantly higher than those in control subjects.12 Extensive studies have confirmed that MMP expression and activity of MMP1, 2, 3, 9, and 12 are associated with asthma pathogenesis and asthma-associated airway remodeling.9 Using animal models, an increase in MMP9 activity in airway mucosa was shown to be associated with epithelial damage, subepithelial basement membrane alterations, and subepithelial collagen deposition.13 Additionally, epithelium-derived metalloproteinases were shown to be involved in the pathogenesis of asthma.14
The airway epithelium is the first barrier to respond to environmental triggers (e.g., pollution, viral infection, and allergens) and is pivotal in asthma pathogenesis.15 Under physiological conditions, the epithelium forms a highly regulated and impermeable tight junction-containing barrier. Tight junctions comprise transmembrane proteins (e.g., occludin, claudins, and junctional adhesion molecules) and peripheral membrane proteins [e.g., zonula occludens (ZO), cingulin], which interact to form a complex protein network.16 Several reports demonstrated that the barrier function of the airway epithelium is impaired in asthma, which enables allergen particles to penetrate the airway wall, inducing immune and inflammatory responses and causing tissue damage.1718 Furthermore, MMP9 has been shown to modulate tight junction integrity and cell viability directly in the human airway epithelium.19
In this study, we demonstrated that German cockroach extract (GCE) induces MMP1 increase in NCI-H292 cells, which directly influences tight junction change. Furthermore, we discovered that the MMP1-induced tight junction alteration is controlled by transcription factors (ETS1 and SP1) activation and extracellular signal-regulated kinase (ERK)/mitogen-activated protein kinase (MAPK) pathway. In addition, we noted that TLR2 might be involved in MMP1-induced tight junction disruption.
MATERIALS AND METHODS
Reagents
GCE was purchased from the Arthropods of Medical Importance Resource Bank, Yonsei University College of Medicine (Seoul, Korea). Small interfering RNAs (siRNAs) against AP1, NF-κB, SP1, ETS1, and MMP1 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-TLR2 antibody and anti-phosphorylated and control p44/42 antibodies were purchased from R&D Systems (Minneapolis, MN, USA) and Cell Signaling Technology (Denvers, MA, USA), respectively. The broad-spectrum MMP inhibitor GM6001 and the MAPK/ERK inhibitor PD98059 were purchased from Santa Cruz Biotechnology and Cell Signaling Technology, respectively.
Cell culture
Human mucoepidermoid pulmonary carcinoma cells (NCI-H292) (ATCC, Manassas, VA, USA) were plated into six- or 12-well culture plates in RPMI 1640 supplemented with 10% fetal bovine serum and 100 U/mL penicillin and streptomycin (GE Healthcare Life Sciences, Logan, UT, USA). Their morphology is epithelial and culture properties are adherent (ATCC).
Real-time PCR
Total RNA was isolated from cells using TRIzol reagent, and cDNA was synthesized using random primers and SuperScript III reverse transcriptase (Invitrogen, Carlsbad, CA, USA). Real-time PCR was performed with AccuPower GreenStar qPCR premix using ExicyclerTM 96 Real-Time Quantitative Thermal Block (Bioneer, Seoul, Korea), and mRNA expression levels were normalized to those of the housekeeping gene GAPDH. Primers are shown in Supplementary Table 1 (only online).
ELISA
Human MMP1 and 9 protein levels in culture cell supernatants were determined using ELISA kits following the manufacturer's instructions (R&D Systems).
Western blotting
Following stimulation with GCE, cells were washed with cold phosphate-buffered saline (PBS) and lysed using M-PER supplemented with proteases (Thermo Fisher Scientific, Waltham, MA, USA). Cell lysates were centrifuged at 12000 rpm for 5 min, and the total protein contents were analyzed using a Bradford assay (Bio-Rad, Hercules, CA, USA). Protein extracts were subjected to 3.5–10% SDS-PAGE and Western blot analysis with anti-ZO-1 and anti-occludin antibodies (Invitrogen). Proteinantibody complexes were observed using the X-ray films and the ECL detection kit (GE Healthcare, Amersham, UK).
Immunofluorescence
Following exposure to GCE, cells seeded onto the Nunclon Delta Surface (Thermo Fisher Scientific) were washed with PBS, fixed in 4% paraformaldehyde for 20 min at room temperature, incubated successively in 1 mM NH4Cl and 4% Tween, labeled with a primary antibody against ZO-1 and occludin for 1 h, and incubated with FITC-labeled goat-anti-mouse IgG conjugate (Invitrogen). Cell-antibody complexes were analyzed on an Olympus Fluo FV100 confocal microscope (Olympus Corporation, Tokyo, Japan).
Electrical resistance
Transepithelial electrical conductance was monitored dosedependently using a volt-ohm meter and chopstick electrodes, according to manufacturer's instructions (World Precision Instruments, Sarasota, FL, USA). PBS was applied for each measurement and then removed.
siRNA transfection
Cells were transiently transfected with the siRNAs against NF-κB, AP1, SP1, and ETS1 and siRNA against MMP1 using the TransIT-X2 Dynamic Delivery System (Mirus Bio, Madison, WI, USA) and were incubated in complete media overnight, following the manufacturer's recommendations. After the GCE treatment, the supernatants were harvested for MMP1 ELISA, while the cells were prepared for MMP-1 qRT-PCR and Western blot analysis of tight junction proteins.
Preparation of nuclear extracts and EMSA
Nuclear extracts from GCE-treated cells were prepared using NE-PER nuclear and cytoplasmic extraction reagents (Pierce, Rockford, IL, USA). Oligomers encoding the consensus sequences of NF-κB, AP1, SP1, and ETS1 were labeled with biotin using a DNA 3' End Biotinylation kit. For the binding reactions, 3 µg of nuclear protein extracts were incubated with binding buffer, poly (dI-dC), and biotin-labeled probe in a total volume of 20 µL for 20 min at 25℃, according to the manufacturer's instructions (Pierce). The protein-DNA complexes were separated on a 6% polyacrylamide gel and transferred to a nylon membrane. The membrane was UV cross-linked, and the biotin-labeled DNA molecules were detected by chemiluminescence (Pierce).
RESULTS
GCE induces MMP expression in airway epithelial cells
To investigate whether GCE can induce MMP expression, since some MMPs are found to be remarkably increased in asthmatic patients,12 we stimulated human airway epithelial cells (NCI-H292) with GCE (50 µg/mL) for 0–48 h. Afterwards, cells were harvested to determine MMP1, 2, 7, 9, 12, and TIMP1 mRNA expression, while cell culture supernatants were utilized to determine MMP protein levels using ELISA. As shown in Fig. 1A, GCE significantly induced only MMP1 (9.9-fold) and MMP9 (18.7-fold) mRNA expression at 6 h, compared with the GCE untreated cells as the control (n=3, p<0.05). Timecourse expression analyses showed that MMP1 significantly increased at 12 h after initial GCE treatment (23.2-fold, compared to that at 0 h), while MMP9 expression peaked at 6 h and gradually decreased thereafter (Fig. 1B and C). Furthermore, as TIMP is known to be one of the important endogenous inhibitors of MMPs,12 we analyzed the expression of TIMP1 as well. However, no correlation among MMP1, 9, and TIMP1 expression following the GCE treatment was observed (Fig. 1D). Moreover, MMP1 and MMP9 protein levels in the supernatant (99.9±3.7 pg/mL to 1010.1±96.1 pg/mL and 10.3±0.7 pg/mL to 58.0±1.8 pg/mL, respectively) at 6 h and 48 h after GCE treatment were shown to be significantly higher than those the control (n=3, p<0.05) (Fig. 1E and F). However, as the MMP1 expression was shown to be much higher than MMP9, we continued our research focusing on MMP1.
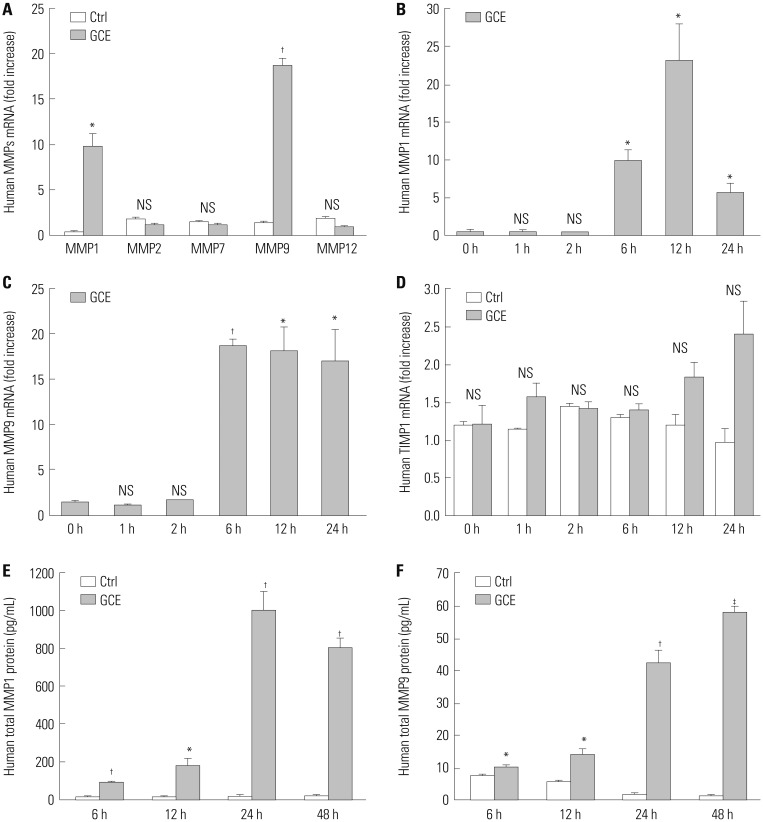
Fig. 1
Expression of matrix metalloproteinases (MMPs) and TIMP in human airway epithelial cells (NCI-H292) treated with German cockroach extract (GCE) for up to 48 h. (A) MMP1, 2, 7, 9, and 12 mRNA expressions at 6 h after GCE treatment. Time-dependent mRNA expression of (B) MMP1, (C) MMP9, and (D) TIMP1 was analyzed using real-time PCR. (E) MMP1 and (F) MMP9 protein expression were analyzed in the cell culture supernatants. Data are presented as the mean±SEM obtained in three independent experiments. *p<0.05, †p<0.01, ‡p<0.001, compared with control (ctrl, media only) or cells at 0 h of media only. NS, not significant.
![]()
GCE leads to disruption of the cellular tight junction
We treated cells with GCE for 24 h and stained them with anti-occludin or anti-ZO-1 antibody, since these proteins are important indicators for the disruption of the cellular tight junction: Occludin plays a crucial role in the maintenance of tight junctions and junction remodeling, while ZO-1 is involved in the establishment of the belt-like tight junctions. In addition, these proteins are known to closely interact, although the characteristics and functional significance of the interaction are poorly understood.16 As shown in Fig. 2A, control cells (untreated) showed strong immunofluorescent staining, with clear intercellular borders, while the GCE-stimulated cells displayed irregular and smeared staining patterns. Consistent with tight junction disruption, GCE treatment significantly and dose-dependently decreased TEER (Fig. 2B). Meanwhile, occludin and ZO-1 mRNA expression did not significantly differ between controls and GCE-treated cells (Fig. 2C and D), whereas occludin and ZO-1 protein expression decreased depending on the dose of GCE treatment (Fig. 2E and F).
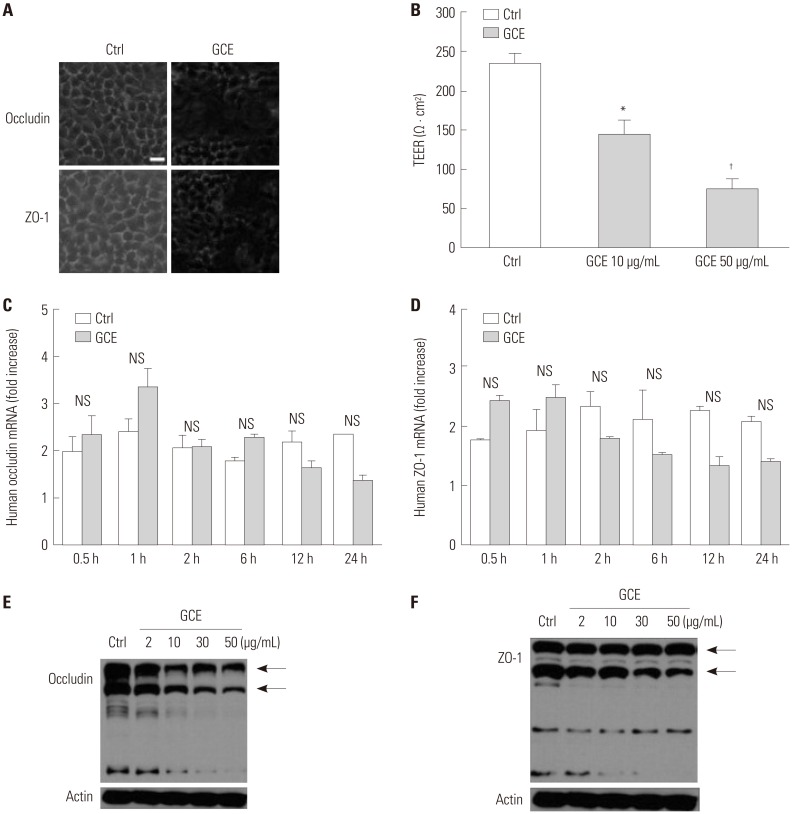
Fig. 2
GCE induces tight junction and its proteins change. (A) Occludin and ZO-1 localization on GCE-stimulated cells was detected by immunofluorescence staining with anti-occludin or ZO-1 and observed under confocal microscopy. Scale bar represents 10 µm. (B) Transepithelial electrical resistance was measured after 24 hours of GCE treatment. (C) Occludin and (D) ZO-1 mRNA expression by time-dependent GCE treatment. *p<05 and †p<0.01, compared with control (ctrl, cells in media only). (E) Occludin and (F) ZO-1 protein expression by dose-dependent GCE treatment probed with anti-occludin or ZO-1. Control (Ctrl) is protein from cells in media only probed with anti-occludin or ZO-1. Data are presented as the mean±SEM obtained in three independent experiments. Representative images are presented. GCE, German cockroach extract; ZO, zonula occludens; NS, not significant.
![]()
GCE-induced MMP1 expression leads to tight junction disruption
To determine the relationship between tight junction disruption and MMP1 release, cells were pretreated with an MMP inhibitor, GM6001, that can inactivate MMP1, 2, 3, 7, 8, 9, 12, 14, and 26 for 1 h prior to the 24-h treatment with GCE. Confocal microscopy analysis showed that GCE-treated cells had discontinuous tight junctions, together with the altered localization of occludin and ZO-1, whereas GM6001 prevented the cellular junctions from being disrupted (Fig. 3A). Furthermore, we determine that MMP1 siRNA transfected cells maintained occludin and ZO-1 protein expression even after GCE treatment via immunoblotting analysis (Fig. 3B and C). The transfection efficiency of MMP-1 siRNA was confirmed by the measurement of MMP1 protein in the supernatant (Fig. 3D)
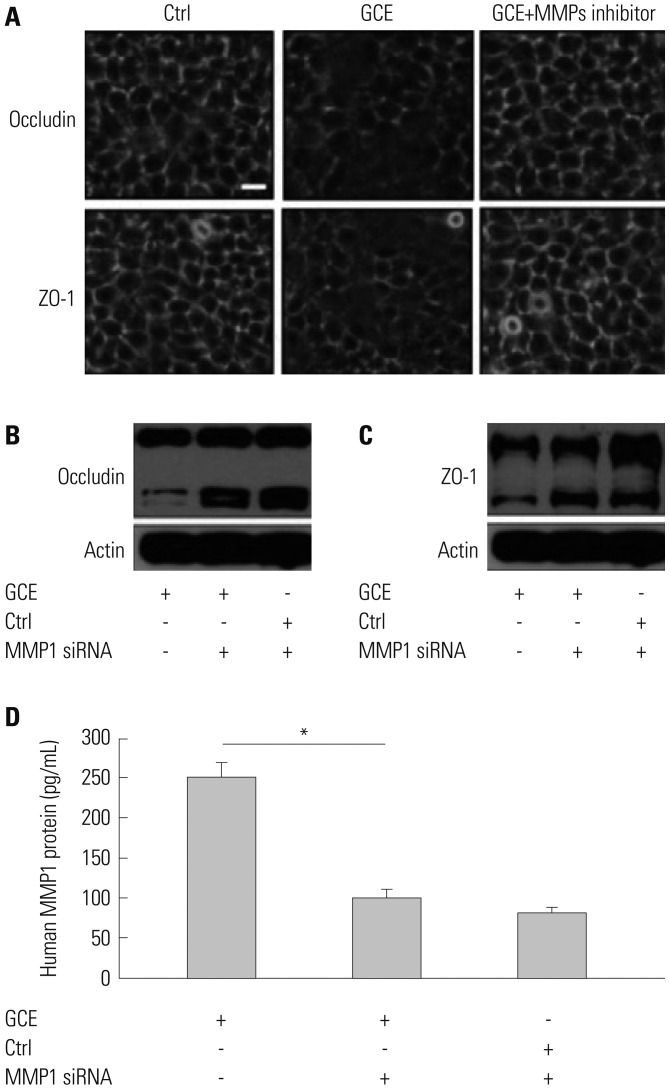
Fig. 3
GCE-induced MMP1 expression influences tight junction disruption. (A) Representative images show occludin and ZO-1 localization in cells pretreated with MMP inhibitor GM6001 and treated with GCE. Scale bar represents 10 µm. (B) Occludin and (C) ZO-1 protein was detected in cells transiently transfected with MMP1 siRNA and treated with GCE by anti-occludin or ZO-1. Representative images of three independent experiments are presented. (D) MMP1 protein expression abundance was measured in the supernatant of cells transiently transfected with MMP1 siRNA and treated with GCE by ELISA. *p<05, compared with the GCE only-treated cells. GCE, German cockroach extract; MMP, matrix metalloproteinase; ZO, zonula occludens; siRNA, small interfering RNA.
![]()
ETS1 and SP1 regulate GCE-induced MMP-1 expression that leads to tight junction disruption
Since it was reported that MMP1 expression and dysregulation correlates with asthma severity relating to pulmonary architecture remodeling and inflammation,20 we investigated the mechanisms by which GCE regulates MMP1 expression. First, we analyzed mRNA expressions of transcriptional factors, ETS1, SP1, AP1, and NF-κB, known to play important roles in MMP1 regulation.2122 As shown in Fig. 4A and B, ETS1 and SP1 mRNA expression was highest at 6 h and 1 h, respectively, after GCE treatment (2.7-fold and 1.5-fold, respectively, p<0.01, compared with GCE untreated cells as the control; n=3). However, AP1 and NF-κB mRNA expression did not change (data not shown). Through EMSA analysis, we confirmed that ETS1 abundantly translocated into the nucleus at 1 h after GCE treatment, while SP1 did so at 2 h (Fig. 4C and D). Second, we evaluated whether ETS1 and SP1 can regulate GCE-induced MMP1 expression using siRNA transfection. The results indicated that ETS1, ETS2, and SP1 siRNA inhibit GCE-induced MMP1 expression (74.4, 75.2, and 60.7% reduction, respectively, compared with the GCE-stimulated cells; n=3) (Fig. 4E and F). Furthermore, ETS1 and SP1 siRNA transfected cells were treated with GCE for 24 h and lysed for immunoblotting analysis to investigate tight junction protein expression. It was previously confirmed that cells transiently transfected with ETS1 and SP1 siRNA in media only show no difference in occludin and ZO-1 protein probed with anti-occludin and ZO-1 (Supplementary Fig. 1, only online). In the result, the occludin protein expression did not significantly change against siRNA ETS1 and SP1 transfection after GCE treatment, while ZO-1 protein expressions in ETS1 and SP1 siRNA transfected cells were not influenced by GCE (Fig. 4G and H).
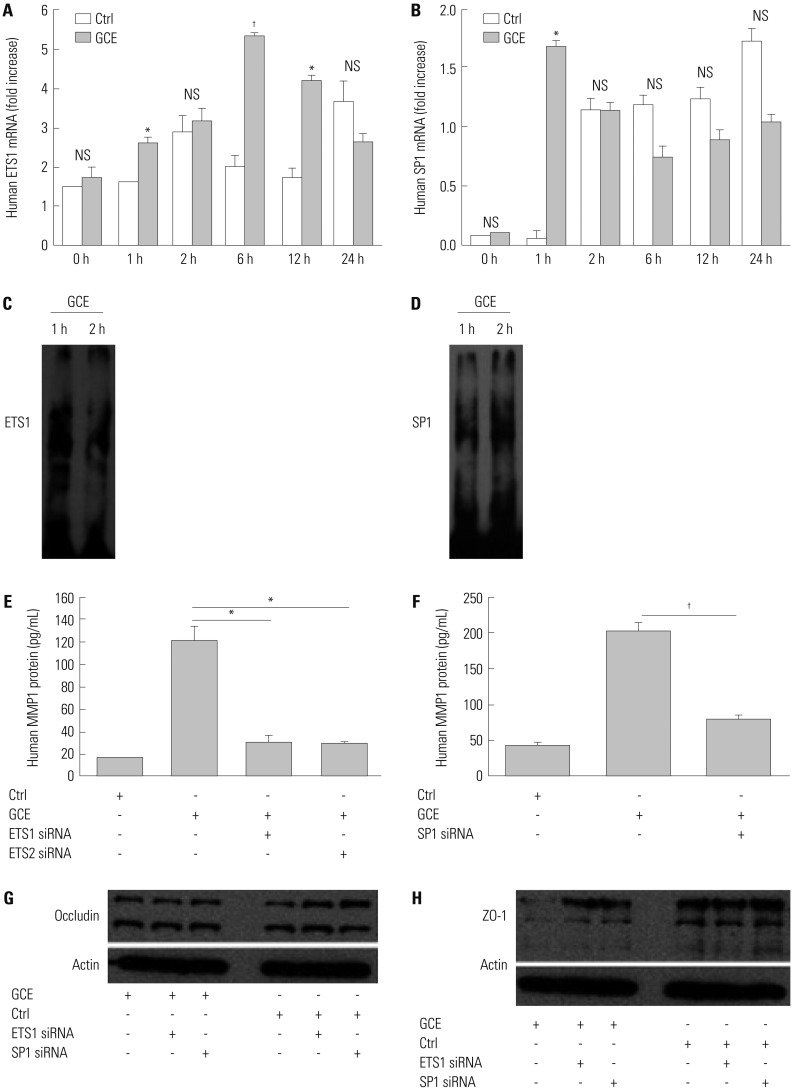
Fig. 4
GCE-induced MMP1 transcriptional regulation influences tight junction disruption. (A) ETS1 and (B) SP1 mRNA was expressed in GCE-treated cells. (C and D) Nuclear extracts were collected to assess the translocation of ETS1 and SP1. (E and F) MMP1 protein was measured in transiently transfected cells with ETS1, ETS2, or SP1 siRNA and treated with GCE. (G and H) Cell lysates were collected to assess the expression of occludin and ZO-1 protein. Data are represented as the mean±SEM obtained in three independent experiments. *p<05, †p<0.01, compared with cells in media only or treated with GCE. Images are representative of three individual experiments. GCE, German cockroach extract; MMP, matrix metalloproteinase; ZO, zonula occludens; siRNA, small interfering RNA; NS, not significant.
![]()
ERK through TLR2 controls GCE-induced MMP-1 expression, leading to tight junction protein decrease
Furthermore, we investigated GCE-induced MAPK pathway activation using specific antibodies against phosphorylated ERK1/2, p38, and JNK (data not shown). While GCE activated the phosphorylation of ERK1/2 molecules (Fig. 5A), it did not activate p38 and JNK molecules (data not shown). Cell treatment with PD98059, a potent inhibitor of MAPK/ERK, significantly decreased GCE-induced MMP1 expression (n=3; 65.9% reduction, compared with that in the GCE only-treated cells; p<0.05) (Fig. 5B). Furthermore, this treatment prevented the GCE-induced translocation of ETS1 and SP1 (Fig. 5C and D). While PD98059 inhibitor did not affect GCE-induced occludin decrease, it did prevent GCE-induced decreases in ZO-1 levels (Fig. 5E and F). Additionally, we investigated whether TLRs, which play a key role in innate immune system responses, can affect GCE-induced MMP1 release, leading to tight junction alterations. A specific anti-TLR2 antibody prevented GCE-mediated MMP1 expression (97% of reduction, compared with that in the GCE only-treated cells; n=3; p<0.01) (Fig. 6A) and GCE-induced decrease of occludin protein expression as well (Fig. 6B). However, ZO-1 protein expression was not affected by anti-TLR antibody pretreatment (Fig. 6C).
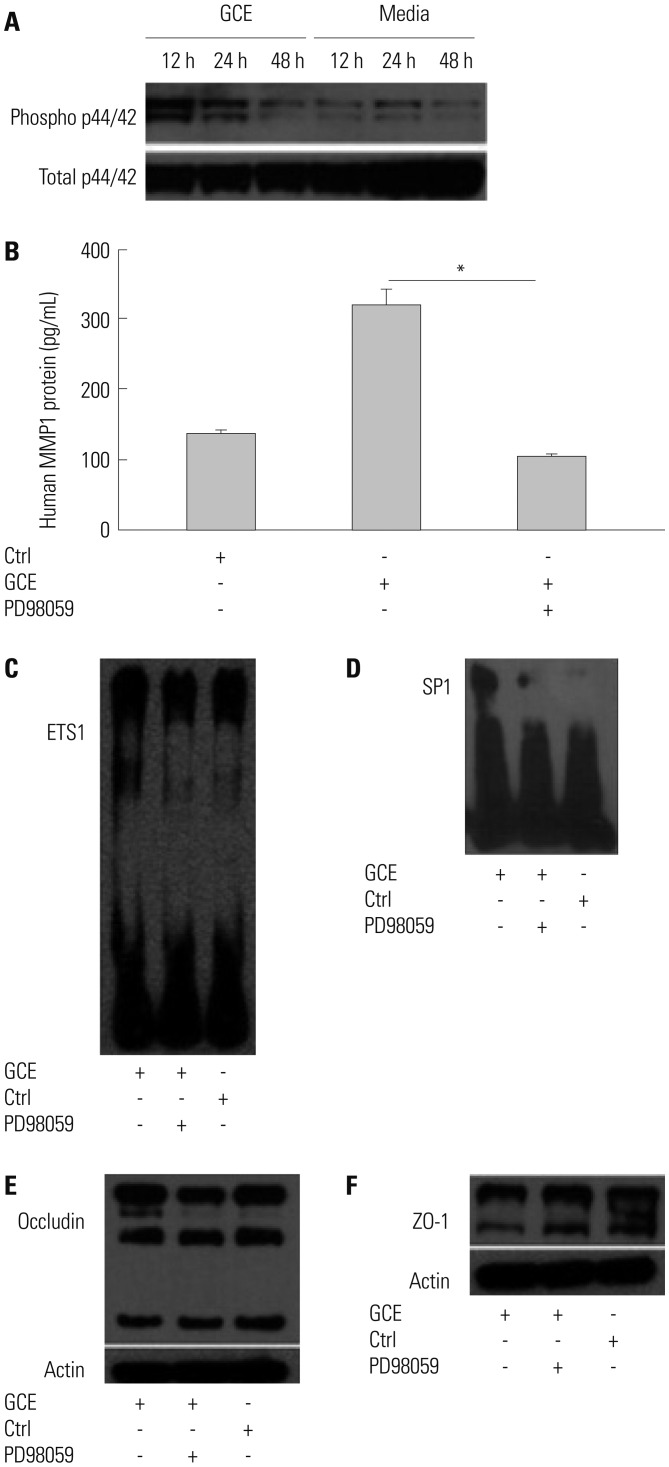
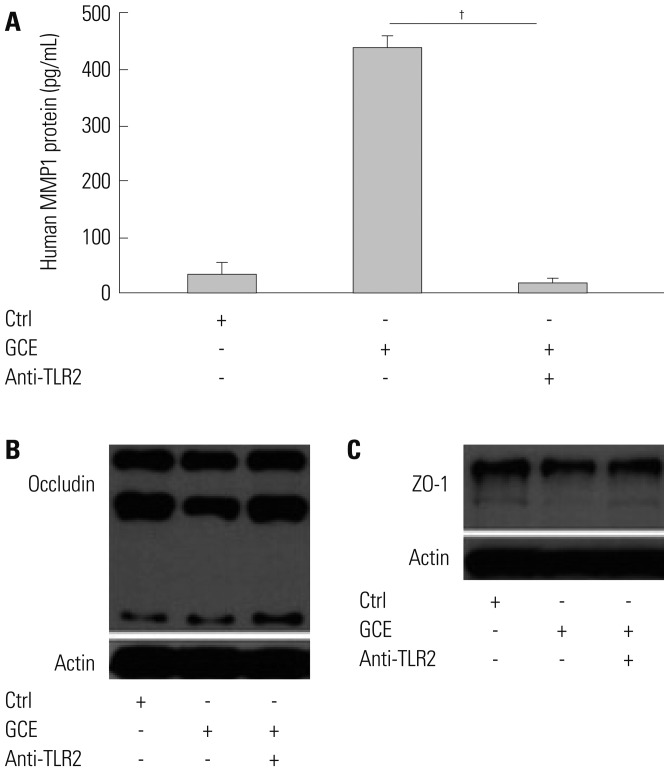
Fig. 5
ERK/MAPK pathway modulates MMP1 transcriptional regulation and tight junction disruption. (A) Phosphorylated and total ERK1/2 was detected in H292 cells pretreated with PD98059 for 1 h and treated with GCE for 24 h. (B) MMP1 protein was measured in the supernatant. Data are presented as the mean±SEM obtained in three independent experiments. *p<05, compared with the GCE only-stimulated cells. (C) ETS1 and (D) SP1 translocation were identified by the biotin-labeled probe of ETS1 or SP1. (E and F) Occludin and ZO-1 protein were immunoblotted with anti-occludin and ZO-1 in cells pretreated with PD98059 and treated with GCE. Images are representative of the results obtained in three independent experiments. MMP, matrix metalloproteinase; GCE, German cockroach extract; ZO, zonula occludens; ERK, extracellular signal-regulated kinase; MAPK, mitogen-activated protein kinase.
![]()
Fig. 6
TLR2 regulates MMP1 expression and might affect tight junction disruption. (A) MMP1 was measured in cells pretreated with anti-TLR2 antibody and treated GCE. Data are presented as the mean±SEM obtained in three independent experiments. †p<0.01, compared with the GCE only-treated cells. (B and C) Occludin and ZO-1 protein was detected in cells pretreated with anti-TLR2 and treated GCE by anti-occludin and ZO-1 antibody. Images are representative of the results obtained in three independent experiments. TLR2, Toll-like receptor 2; MMP, matrix metalloproteinase; GCE, German cockroach extract; ZO, zonula occludens.
![]()
DISCUSSION
Cockroach allergen was shown to contain a protease that associates with PAR2 and TLR2 motifs, which can modulate inflammatory cytokine and chemokine activity, leading to direct activation of the innate immune system.57 Furthermore, cockroach extract increases bronchial airway epithelial permeability, although the roles of different components have not been elucidated.23 Here, we demonstrated in vitro that GCE significantly induces MMP1 expression. MMPs are key molecules involved in cell proliferation and migration, tissue growth, remodeling, and regeneration and have high proteolytic activity and broad substrate specificity.10 Recently, MMPs were identified as likely to be involved in the pathogenesis of asthma, primarily in the asthma-associated airway remodeling.24 Although the role of MMP9 in asthma pathogenesis has been extensively studied, the levels of other MMPs, including MMP-1, 2, 3, or 12, in individuals with asthma were shown to be increased as well, indicating that their levels in sputum, BAL, and exhaled breath condensates correlate with disease exacerbation.2526 Additionally, it was reported that MMP1 is highly expressed in airway epithelial cells, inflammatory cells, and even airway smooth muscle cells in asthmatic patients.202728
Many studies have indicated that various aeroallergens, such as house dust mite and pollen, exhibit proteolytic activities, which may contribute to asthma pathogenesis by damaging the barrier function of the airway epithelium.2930 However, only a small number of studies have investigated the effects of cockroach allergen on the epithelial barrier.5 In this study, we selected occludin and ZO-1 as the indicators of tight junction disruption, because occludin is a key component of a tight seal formed between the cells and because ZO-1 directly interacts with occludin to ensure proper localization of the tight junctions.16 We showed that GCE induces morphological changes and a GCE dose-dependent reduction of tight junction protein levels, in combination with an increase in the epithelial permeability, suggesting that GCE may lead to the degradation of epithelial tight junction proteins. Previously, cockroach antigen treatment was shown to induce VEGF expression and decrease electrical resistance in bronchial epithelial cells, although the cockroach role was not investigated.23 To date, the activity of cockroach allergen has been explained by its effects on PAR2, which can lead to disease pathogenesis or exacerbation.5 It was also suggested that these cockroach allergen activities mediated by PAR2 may be further complicated by the involvement of TLRs, showing that cockroach allergen contains both TLR4 and TLR2 motifs7 and that PAR2 and TLR4 signal cooperatively.3132
Our data demonstrated that GCE induces MMP1 expression and tight junction disruption, which play important roles in disease-associated airway remodeling.19 Therefore, we investigated whether MMP1 cleaves occludin and ZO-1, the components of tight junctions. In the result, GM6001, a potent MMP inhibitor, and MMP1 siRNA prevented the degradation of tight junction proteins, showing that tight junction disruption may be increased by GCE-induced MMP1 expression in airway epithelial cells. MMP1 is a collagenase that can be detected in the small airways and lung parenchyma of asthmatic patients.3334 MMP1 expression was shown to directly correlate with the airway obstruction in BAL fluid and to be associated with the development of asthma symptoms, although the mechanisms thereof remain unclear.202835 Inadequate tight junction formation has been confirmed in asthmatic samples, compared with normal cultures, using anti-ZO-1 and anti-occludin antibodies, suggesting that respiratory viruses, air pollutants, and proteolytically active allergens can cause epithelial desquamation through tight junction disruption and the disruption of epithelial barrier function.1820293036
Although the role of MMPs in asthma pathogenesis has been extensively researched, their inhibitors for potential therapeutic purposes have rarely been developed. Therefore, understanding the molecular mechanisms underlying the regulation of MMP1 expression may help identify potential targets for the prevention of disease development. MMP1 expression is tightly regulated at the levels of transcription, post-transcription, and post-translation.37 At the transcriptional level, the promoter of MMP1, which contains adjacent binding sites for ETS1 and AP1 transcription factors, is known to be crucial for the regulation of MMP1 expression.2138 The distal 1.5-kb segment of the MMP1 promoter was identified as necessary for the direct induction of MMP1 expression by cigarette smoke.39 Moreover, NF-κB is a specific transcription factor that localizes to the promoter regions of MMPs in response to various stimuli.2122 Here, we analyzed the involvement of these transcription factors, ETS1, AP1, SP1, and NF-κB, in GCE-induced MMP1 expression, showing that both ETS1 and SP1 are significantly activated and that their inhibition attenuates MMP1 release and ZO-1 degradation. Furthermore, we investigated the signaling molecules mediating GCE-induced MMP1 release, and the TLR2 and ERK/MAPK pathway activation were shown to be required for the increase in MMP1 levels. Further, ERK/MAPK signaling molecules were shown to regulate ETS1 and SP1 translocation induced by GCE, following TLR2 activation. MMP1 inhibition prevented tight junction degradation. Although TLRs are expressed on the majority of cells, they alone are not sufficient for the recognition of pathogens without coreceptors and adaptor molecules.7 The role of TLR2 in asthma is controversial since different studies have shown that it can act as an enhancer or inhibitor of the allergy-related airway inflammation during sensitization.40 German cockroach frass was suggested to contain a TLR2 agonist that directly induces neutrophils infiltration and further exacerbation of airway inflammation.7 Furthermore, it was reported that Staphylococcus aureus induces MMP1 expression in human bronchial epithelial cells through the activation of TLR2/EGF receptor signaling axis.38 The AP1, NF-κB, SP1, and ETS transcription factors and MAPK family members were shown to play crucial roles in the MMP induction. MMP1 expression induced by one of the extracellular matrix proteins, tenascin-C, was shown to depend on ERK1/2, JNK, and p38 MAPK activation in airway smooth muscle cells.20 Cockroach frass-induced MMP9 expression can be upregulated by ERK activation and AP1 nuclear translocation.5
In conclusion, we demonstrated in this study that GCE induces an increase in MMP1 release, known to be a pro-remodeling factor and to play a crucial role in tight junction disruption in airway epithelial cells. In order to assist the future development of an MMP1 inhibitor, we identified transcription factors, such as ETS1 and SP1, and signaling molecules, such as TLR2 and ERK/MAPK, as regulating MMP1 expression. These molecules may represent potential targets for the development of novel treatments for airway diseases such as asthma.
 XML Download
XML Download