

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Hepatocellular carcinoma (HCC) is the fifth leading cause of cancer mortality worldwide.12 The main cause of HCC is cirrhosis, which originates from infections caused by chronic hepatitis B virus, hepatitis C virus, or alcohol consumption.34 Patients with early-stage HCC are often asymptomatic, and it is usually detected at intermediate or advanced stages in which patients cannot be treated by curative hepatic resection or liver transplantation.5 In addition, although surgical treatment for early HCC has improved patient outcome, the risk of recurrence remains substantial and there is still no curative therapy for advanced HCC. Therefore, there is an increasing need for effective early diagnosis and development of novel therapeutics for HCC patients. Owing to its important role in metabolic reprogramming during carcinogenesis, cancer metabolism has gained popularity in the fields of cancer diagnosis and therapy. This review summarizes the current state of research related to cancer metabolism, to help identify potential new therapeutic targets for HCC.
CLINICAL APPLICATIONS OF METABOLIC IMAGING IN HCC
Alpha-fetoprotein (AFP) was the first glycoprotein identified as a marker of HCC, and this protein is used to screen for this particular disease.6 However, approximately 30% of HCC patients maintain normal AFP levels, and some HCC patients also have relatively low levels of AFP.7 To overcome these problems, several imaging techniques are used to diagnose patients with suspected HCC, such as ultrasound, computed tomography (CT), magnetic resonance imaging (MRI), positron emission tomography/computed tomography (PET/CT), and angiography.
Of the imaging modalities, clinical efficacy of functional imaging based on cancer metabolism for the assessment of HCC has been actively investigated. F-18 fluorodeoxyglucose (18F-FDG), a surrogate for enhanced glucose metabolism, has been used widely for HCC. The uptake mechanism and biochemical pathway of 18F-FDG metabolism has been extensively studied in vitro and in vivo; transport through cell membrane via glucose transporter isoform 1 (GLUT1) and intracellular phosphorylation by hexokinase (HK) have been identified as key steps for subsequent accumulation in HCC.8910111213 Many studies have shown upregulation of GLUT1 in HCC, but not in non-tumor liver tissue.1415 Amann, et al.15 demonstrated a positive correlation between GLUT1 expression and Ki-67 labeling index in patients with HCC, suggesting that its expression is associated with advanced tumor stage and poor differentiation. In addition, poor survival has been reported in patients with high tumor GLUT1 expression, based on The Cancer Genome Atlas (TCGA) data set.16 A relationship between enhanced FDG uptake and dysregulation of epithelial-mesenchymal transition-related proteins was demonstrated in HCC through in vitro and patient tissue experiments.16 Representative of the underlying biological characteristics of tumor, 18F-FDG PET/CT images are predictive of tumor recurrence or survival after various treatments.17
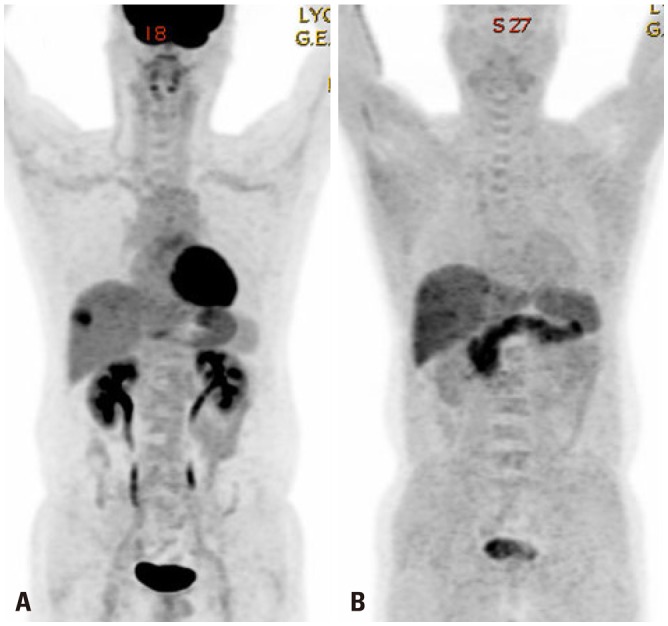
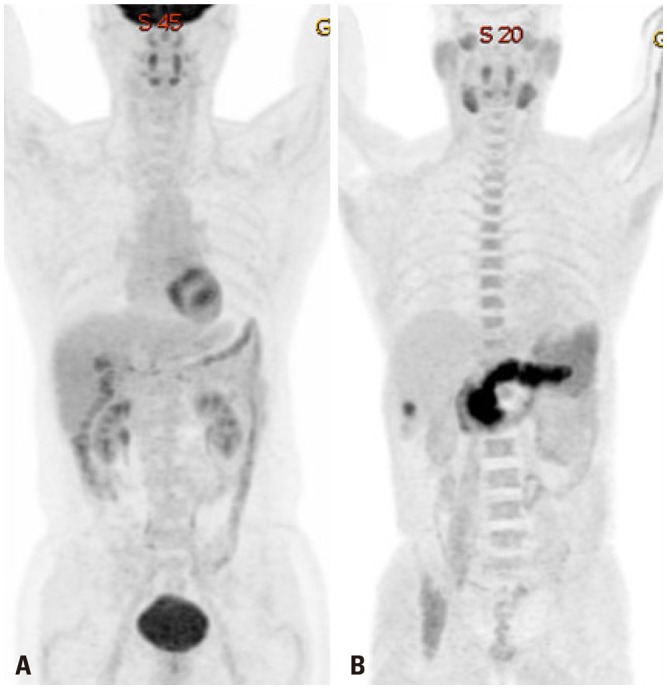
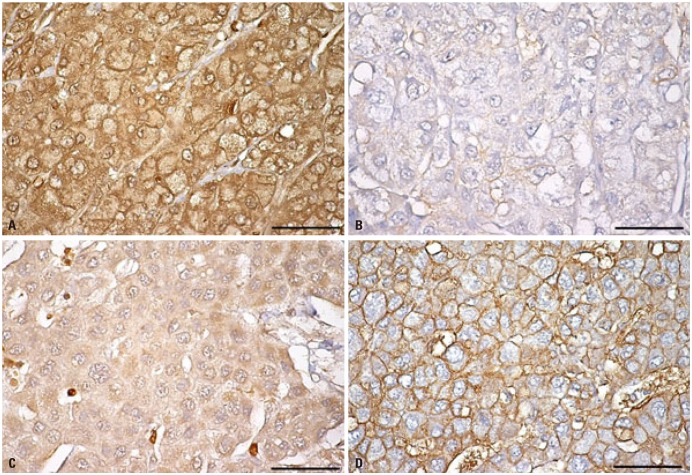
Despite displaying increased glycolysis even with the presence of oxygen, the so-called Warburg effect, HCCs are notorious for exhibiting a wide spectrum of 18F-FDG uptake capabilities, considerably reducing the sensitivity of tumor detection. Alternatively, C-11 acetate has been proposed as a radiotracer for detecting tumors that are not identified based on 18F-FDG uptake (Figs. 1 and 2). Acetate is a source of acetyl-CoA, and it plays an essential role in regulating the activity and expression of proteins involved in regulation of intracellular biomass, lipogenesis, and acetylation.18 Acetate was shown to be utilized by tumors as an alternative nutrient under low cellular glucose uptake conditions, and C-11 acetate accumulation in tumors has been found to be associated with tumor progression.19 HCC has been reported to use acetate as a substrate for fatty acid biosynthesis through up-regulation of acetyl-CoA synthase and monocarboxylate transporter (MCT).20 Recent studies have indicated that MCT1 is a novel import system of acetate in non-glycolytic HCC tumors. Indeed, Fig. 3 shows various expressions of GLUT1 and MCT1 in HCC patients with different levels of 18F-FDG and 11C-acetate uptake. It was demonstrated that absorption of acetate by MCT1 promotes oxidative phosphorylation and lipid metabolism in non-glycolytic HCC tumors.21 Accordingly, combining 18F-FDG PET/CT with C-11 acetate PET/CT could be useful to provide relevant information on prognostic and molecular heterogeneity.
DIFFERENTIAL GENE EXPRESSION THAT ALTERS METABOLISM IN HCC CELLS
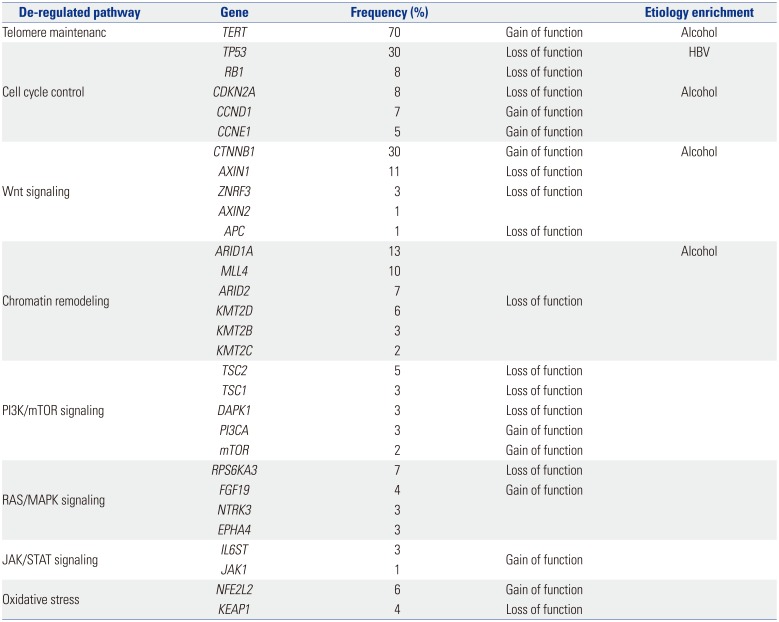
HCC is a heterogeneous disease, both clinically and from a molecular standpoint. Different risk factors such as hepatitis virus infection, aflatoxin exposure, or alcohol abuse are linked to specific pathways, and these can be strongly associated with certain types of HCC. Based on the results of HCC tumor sequencing, different driver genes and associated oncogenic pathways have been identified, based on the composition of tumor source.22232425 Therefore, heterogeneity should be investigated to determine the etiological cause and affected pathways of HCC. High levels of heterogeneity are clinically relevant, as they lead to inconsistent treatment outcomes. Recently, deep sequencing/next generation sequencing has provided new insights into the complex molecular pathogenesis of HCC, including the identification of novel oncogenic pathways and driver genes.252627282930313233 Aberrant telomerase reverse transcriptase (TERT) activation is the most common somatic alteration observed in HCC (~70%). In addition to TERT, CTNNB1, TP53, and Axin1 are mutated at high frequency in HCC. Table 1 summarizes the most relevant mutations in HCC.
Warburg effect occurs downstream of survival signaling pathways, which are altered by loss of tumor suppressor genes or activation of oncogenes such as c-Myc, Ras, Akt, TP53, and HIF-1α.3435363738
c-Myc was reported to result in mouse liver tumors with elevated glycolysis39. HIF-1α, a major transcription factor involved in hypoxic response of cancer cells,40 has been shown to play an important role in several cancers by promoting tumorigenesis, and might also be involved in the “metabolic reprogramming” of cancer cells.4142 This activates the transcription of genes encoding angiogenic cytokines and growth factors, such as VEGF and glycolytic enzymes including hexokinase1 (HK1), hexokinase2 (HK2), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and pyruvate kinase (PKM).434445 Moreover, HIF-1α enhances chemoresistance and radioresistance, whereas it suppresses differentiation and apoptosis in HCC.4647 As a result, elevated HIF-1α levels are associated with increased patient mortality and metastasis in various tumors, including HCC.484950
Certain gene mutations or loss-of-heterozygosity events alter metabolism in a HIF-1α-dependent manner. Although mutations in PTEN gene rarely occur in HCC, frequent loss of heterozygosity of PTEN allele has been identified (in 20–30% of HCC patients).5152 The loss of PTEN plays a critical role in HCC progression and patient outcome by increasing HIF-1α synthesis and stability.5354 The modulation of HIF-1α expression by epidermal growth factor/phosphatidylinositol 3-kinase (PI3K)/PTEN/AKT/FRAP pathway has implications in tumor angiogenesis.55 A few studies have shown that HIF-1 can be activated by Ras and membrane type-1 matrix metalloproteinases under normal oxygen conditions in cancer cells, providing new insight into regulation of cancer glycolysis beyond hypoxic condition.5657 Bufalin, a cardiotonic steroid, was shown to suppress tumor invasion and metastasis by targeting HIF-1α via PI3K/AKT/mTOR pathway, and thus has potential for HCC targeted therapy.58
Numerous microRNAs (miRNAs) have been shown to be associated with HCC. Six miRNAs have been consistently reported to be dysregulated in HCC, when compared to their expressions in non-tumorous tissue.596061 For example, miR-122 and miR-199a, which act as tumor suppressors by regulating the expression of cyclin G and components of PAK4/Raf/MEK/ERK pathway, are downregulated in this disease. Conversely, miR-21, miR-221, miR-222, and miR-224, which target various molecules including PTEN, SMAD4, CDKN1B, and CDKN1C, are upregulated in HCC. Some miRNAs that regulate cancer metabolism are also dysregulated in HCC. miR-34a plays a major role in regulation of cellular metabolism by targeting SIRT1, a key NAD-dependent deacetylating enzyme involved in a wide range of metabolic processes including lipid metabolism, glucose metabolism, and expression of other metabolic regulators.6263 This molecule inhibits cellular glycolysis by targeting HK1/2 and glucose-6-phosphate isomerase. miR-23a directly targets the key gluconeogenic enzyme glucose-6-phosphatase catalytic subunit (G6PC), and is significantly upregulated in primary human HCC.
NEW TREATMENTS TARGETING CANCER METABOLISM
Although sorafenib has limited efficacy, it is still the only standard treatment available for advanced HCC with portal vein invasion or extrahepatic spread.6465 However, other molecules are being developed for targeted therapy. Recent studies on metabolic regulation of cancer cell growth and metastasis have been actively performed. 2-deoxy-D-glucose (2-DG), a glucose analogue that is able to suppress glycolysis by competitively inhibiting HK2, has an effect on HCC growth.6667 The combination of conventional therapy and 2-DG has been reported to synergistically inhibit the proliferation of sorafenib-sensitive and sorafenib-resistant HCC cells.68 3-bromopyruvate (3-BP) directly inhibits HK2 activity and glycolysis pathway. In vitro and in vivo studies have demonstrated the anticancer effects of 3-BP on HCC, and consequently, this drug has been approved by the FDA.6970
Facilitative glucose transporters (GLUTs) have emerged as key factors that are required for increasing glucose uptake by cancer cells.1415 Therefore, small molecules targeting GLUT1 will inhibit cancer cell growth or metastasis by reducing glucose uptake. To increase GLUT1 targeting specificity, derivatives of GLUT1 inhibitor, such as fasentin7172 and WZB117,73 have been investigated. Regulation of hypoxia by molecules, including HIF-1, is an attractive potential therapeutic target for HCC as well as other cancers. The HIF-1α mRNA antagonist EZN-2968, a novel inhibitor of hypoxia-induced gene activation, is currently in Phase I trial for HCC patients.
CONCLUSIONS
Metabolic reprogramming is essential for angiogenesis, proliferation, invasion, and metastasis of cancer. It is also associated with de-differentiation, anti-apoptotic properties, and resistance to conventional chemotherapy and radiotherapy. In the future, cancer metabolism would represent an attractive potential therapeutic target. With their development, PET/CT scans combined with various metabolic radiotracers will offer clinical importance in selecting patients who would benefit from novel drugs targeting different pathways related to cancer metabolism
 XML Download
XML Download