

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Citrus fruits are well known for their nutrition and health benefits. Yuzu (Citrus junos Tanaka), a common citrus fruit found in Korea and Japan, is rich in antioxidant phenols, and flavonoids with an anti-inflammatory action, such as limonene and hesperidin123. Hirota et al.4 showed that limonene extracted from citron reduced reactive oxygen species production and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB) activity, indicating that citron has both antioxidant and anti-inflammatory properties4. However, despite the several beneficial effects of citron, no study has investigated its effects on the skin.
Rosacea is a common chronic inflammatory disease of the facial skin5; however, the exact pathogenesis of rosacea remains unclear. Recent studies have suggested that dysregulation of innate immunity and the neurovascular/neuroimmune system induces angiogenesis and inflammation, resulting in rosacea symptoms such as flushing, stinging, and telangiectasis678. Excessive production of LL-37 (an active peptide form of cathelicidin) and kallikrein 5 (KLK5), the predominant serine protease responsible for cleavage of cathelicidin into LL-37, has been reported to play a role in the dysregulation of innate immunity91011. LL-37 production is induced by the vitamin D pathway in human keratinocytes; therefore, ultra violet light acts as a trigger for rosacea12. Transient receptor potential vanilloid 1 (TRPV1), also known as the capsaicin receptor, has been reported to be involved in neurogenic dysregulation. It can be activated by heat, ethanol, or spicy food, all of which are triggers for rosacea131415. Vascular endothelial growth factor (VEGF), which has been reported to enhance angiogenesis and/or lymphangiogenesis in the skin with rosacea lesions, is associated with neurovascular dysfunction1617. Interleukin 33 (IL-33), a pro-inflammatory cytokine and an inducer of Th2-mediated responses, has been reported to enhance VEGF expression in keratinocytes18. In addition, abnormally increased expression of LL-37 may induce VEGF expression in epidermal keratinocytes, which corresponds to the enhanced inflammation and vascular response seen in rosacea19.
Therefore, LL-37, KLK5, TRPV1, and VEGF are considered as therapeutic targets for rosacea. This study was aimed at determining the effects of citron essential oils at different concentrations on rosacea mediators, KLK5, LL-37 (induced by 1α, 25-dihydroxyvitamin D3 [VD3]), VEGF, and TRPV1 induced by IL-33 and LL-37 in normal human epidermal keratinocytes (NHEKs) in vitro and evaluating their potential as adjuvant therapeutic agents for rosacea.
MATERIALS AND METHODS
Citron essential oils
Two types of citron essential oils were used in this study, citron seed essential oil and unripe citron essential oil. Citrons were obtained from Goheung, Jeollanam-do. Citron essential oils were extracted by Hisol Co., Ltd (Namwon, Korea).
Culture and viability of NHEKs
NHEKs were purchased from EpiLife (Cascade Biologics, Portland, OR, USA). The cells were cultured in basal keratinocyte growth media (EpiLife) supplemented with human keratinocyte growth supplement and antibiotics (100 U/ml penicillin and 100 µg/ml streptomycin) in a 5% CO2 incubator. Passages 2–9 were used for all experiments. NHEKs (1×104 cells/well) were seeded in a 96-well plate. After the cells were treated with various concentrations of the essential oils, cell viability was determined using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich, St. Louis, MO, USA) assay according to the manufacturer's instructions20.
Expression and regulation of KLKs and LL-37 in NHEKs induced by VD3
To induce KLK and LL-37 expression, NHEKs at 70% confluence were stimulated with VD3 (Sigma-Aldrich)2122. The expression of KLK5, LL-37, and vitamin D receptor (VDR) was evaluated at times ranging from 0 to 48 h and doses ranging from 10−9 to 10−7 M (1 to 100 nM)2324. Regulation of KLK 5, LL-37, and VDR induced by VD3 was also evaluated after the cells were treated with citron essential oils at various concentrations (0.005%, 0.01%, and 0.02%).
Expression and regulation of VEGF and TRPV1 induced by IL-33 and LL-37 in NHEKs
NHEKs at 70% confluence were exposed to different concentrations of recombinant human IL-33 (100 ng/ml; MACS, Auburn, CA, USA) and LL-37 peptides (0.5 µg/ml; ANYGEN, Jangseong, Korea). The expression of VEGF and TRPV1 was evaluated at times ranging from 0 to 48 h and doses ranging from 0.1 to 10 ng/ml19. Regulation of VEGF and TRPV1 was also evaluated after the cells were treated with citron essential oils at various concentrations (0.005%, 0.01%, and 0.02%).
Semi-quantitative reverse-transcription (RT) polymerase chain reaction (PCR) and real-time PCR
Total messenger RNA (mRNA) was isolated from human keratinocytes by using the RNeasy mini kit (Qiagen, Valencia, CA, USA). cDNA was synthesized using the Omniscript RT kit (Qiagen): 1 µg RNA, 10× buffer, dNTP, oligodT, inhibitor, reverse transcriptase, at 37℃ for 1 h, 93℃ for 5 min. A PCR was performed using the PCR-premixture kit (ELPIS, Daejeon, Korea) according to the manufacturer's instructions. Table 1 shows the primer sequences, PCR conditions, and product sizes. The PCR products were analyzed using 1.5% agarose gel electrophoresis, stained with Sybr Safe DNA gel stain buffer (Invitrogen, Carlsbad, CA, USA), and visualized using a luminescent image analyzer (LAS 3000; Fujifilm, Tokyo, Japan).
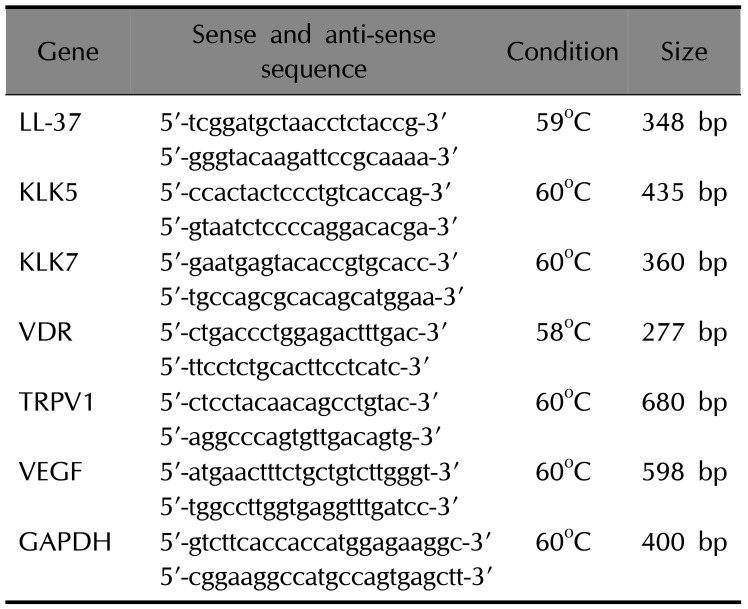
Table 1
Sense and antisense primer sequences, and semi-quantitative PCR and real-time PCR conditions
![]()
Total mRNA was isolated using an RNeasy mini kit (Qiagen). Subsequently, cDNA was reverse transcribed from 500 ng of total RNA with the Omniscript RT kit (Qiagen) and subjected to PCR with the HiPi PCR PreMix (ELPIS). The expression levels were normalized to endogenous glyceraldehyde 3-phosphate dehydrogenase (GAPDH) levels. To determine mRNA levels, RT-PCR and quantitative real-time PCR were performed with the same primer sets for the target genes. Real-time PCR was performed in triplicate with the HOT FIREPol EvaGreen® qPCR Mix Plus (Solis BioDyne, Tartu, Estonia) using a RotorGene 3000 system (Corbett Research, Cambridge, UK). The thermal cycling conditions were as follows: 15 min at 95℃, followed by 40 cycles at 95℃ for 10 s, 55℃–60℃ for 20 s, and 72℃ for 30 s. The relative abundance of a given transcript was estimated using the 2−ΔΔCt method, following normalization to GAPDH levels.
Immunocytofluorescence staining
The cell suspensions were fixed with paraformaldehyde, blocked with 5% goat serum, and incubated with rabbit polyclonal anti-cathelicidin (LL-37) antibody (Abcam, Cambridge, MA USA), mouse monoclonal anti-VEGF antibody (Novusbio Biological, Littleton, CO, USA), or rabbit polyclonal anti-VR1 (TRPV1) antibody (Abcam). AlexFluor 488-conjugated goat anti-rabbit immunoglobulin (Ig) G and AlexaFluor 594-conjugated goat anti-mouse IgG (Molecular Probes; Invitrogen, Carlsbad, CA, USA) were used as secondary antibodies. Sections were mounted using 4′,6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA, USA). The images were visualized using confocal microscopy with a laser scanning microscope (LSM 510; Carl Zeiss, Jena, Germany) and analyzed using the LSM 5 browser imaging software.
Enzyme-linked immunosorbent assay (ELISA)
Commercial ELISA kits were used according to the manufacturers' protocols to quantify the immune molecules of interest: human LL37 (Hycultbiotech, Uden, Netherlands), human KLK5 (R&D Systems, Inc., Minneapolis, MN, USA), human VEGF (R&D Systems, Inc.), and human TRPV1 (Mybiosource, San Diego, CA, USA).
Statistical analysis
All values are expressed as the mean±standard deviation. Statistical analyses were performed using one-way analysis of variance with a post hoc least significant difference test using IBM SPSS Statistics version 21.0 (IBM Co., Armonk, NY, USA) for multiple comparisons. A p-value less than 0.05 was considered statistically significant.
RESULTS
Survival of NHEKs
Cell viability was evaluated after the cells were exposed to various concentrations of citron seed essential oil and unripe citron essential oil (0, 0.001%, 0.002%, 0.004%, 0.008%, 0.016%, 0.031%, 0.063%, 0.125%, 0.25%, 0.5%, and 1%). Viability decreased upon treatment with 1% citron seed essential oil and 0.125% unripe citron essential oil (Supplementary Fig. 1).
Citron essential oils downregulated VD3-induced KLK5 and LL-37 expression in NHEKs
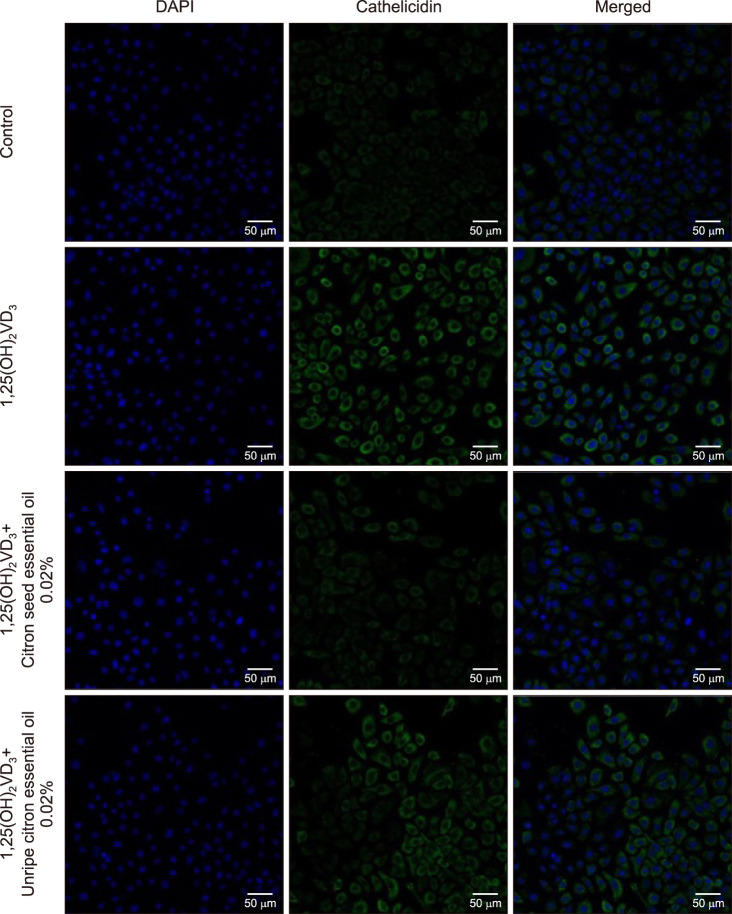
Stimulation of NHEKs with VD3 at a dose of 10−7 M for 24 h increased the levels of KLK5 and LL-37 maximally. After treatment of NHEKs with citron essential oils, KLK5, LL-37 and VDR mRNA expression induced by VD3 decreased. The mRNA levels of LL37 decreased after treatment with citron seed (0.02%) and unripe citron essential oils (≥0.005%), partially showing a dose-dependent pattern (Fig. 1A). Furthermore, the mRNA levels of KLK5 decreased after treatment with unripe citron essential oil (≥0.005%). Citron seed essential oil also decreased the mRNA level of KLK5, but the difference was not significant. Treatment with 0.02% unripe citron essential oil caused the maximum suppression of LL-37 and KLK5 mRNA levels. Further, ELISA showed that protein levels of LL-37 and KLK5 significantly decreased after treatment with citron essential oils at any concentration (Fig. 1B). Immunocytofluorescence showed that LL37 expression induced by VD3 decreased after treatment with 0.02% citron essential oils; this finding was consistent with previous results (Fig. 2).
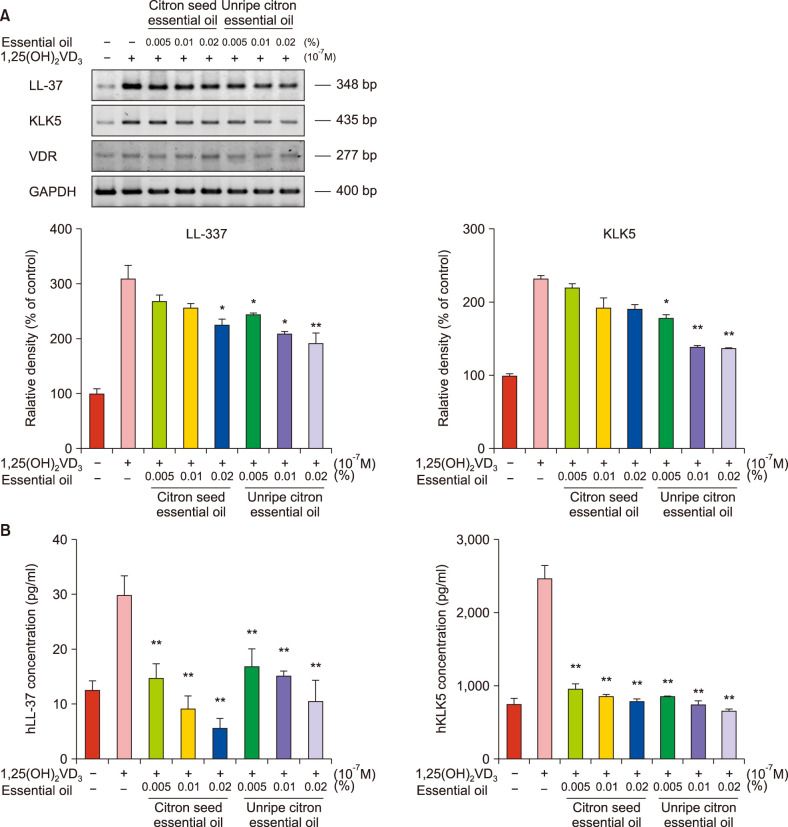
Fig. 1
(A) Messenger RNA levels of kallikrein 5 (KLK5), LL-37, and vitamin D receptor (VDR) induced by vitamin D3 decreased after treatment with citron essential oils in normal human epidermal keratinocytes (NHEKs); semi-quantitative reverse-transcription-polymerase chain reaction. (B) Protein levels of KLK5 and LL-37 induced by VD3 in NHEKs decreased after treatment with citron seed and unripe citron essential oils; enzyme-linked immunosorbent assay. Bars indicate standard deviations. GAPDH: glyceraldehyde 3-phosphate dehydrogenase. *p<0.05, **p<0.01.
![]()
Citron essential oils downregulated VEGF and TRPV1 levels induced by IL-33 and LL-37 in NHEKs
The levels of VEGF and TRPV1 maximally increased upon NHEK exposure to IL-33 100 ng/LL-37 0.5 µg for 24 h. After treatment with citron essential oils, the mRNA levels of VEGF and TRPV1 decreased and the extent of decrease was significant: citron seed essential oil, VEGF ≥0.01% and TRPV1 ≥0.01%, and unripe citron essential oil, VEGF 0.005% and 0.02%, and TRPV1 0.02% (Fig. 3A). Further, ELISA showed that protein levels of VEGF and TRPV1 significantly decreased after treatment with citron seed essential oil (VEGF and TRPV1: 0.02%) and unripe citron essential oil (VEGF and TRPV1: ≥0.01%) (Fig. 3B). Immunocytofluorescence showed that the IL-33-induced TRPV1 and VEGF expression in NHEKs decreased after treatment with 0.02% citron essential oil; this finding was consistent with previous results (Supplementary Fig. 2).
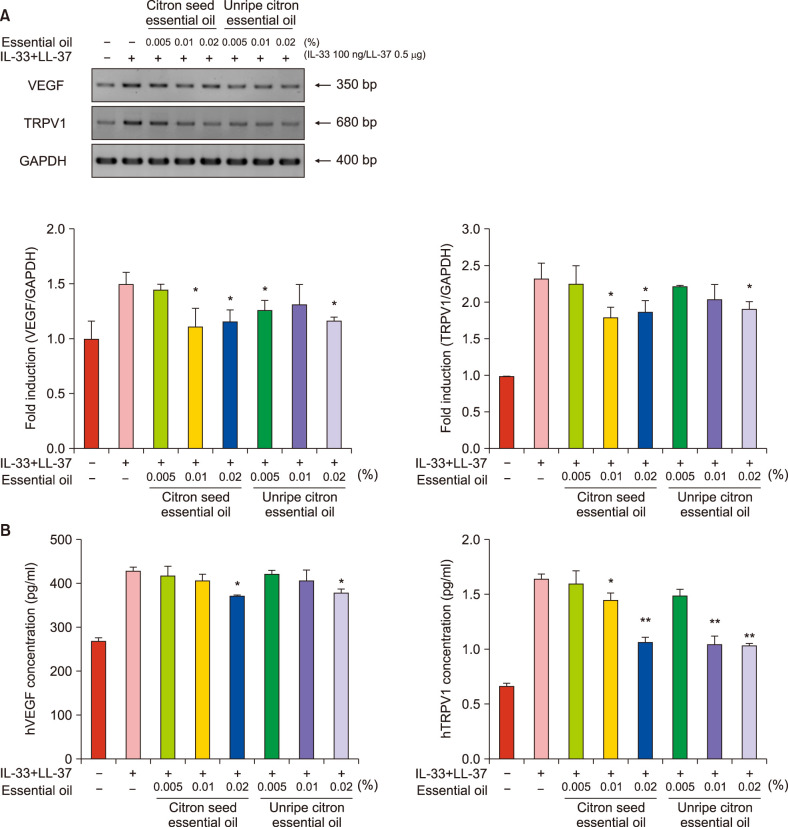
Fig. 3
(A) Messenger RNA levels of vascular endothelial growth factor (VEGF) and transient receptor potential vanilloid 1 (TRPV1) induced by interleukin 33 (IL-33) and LL-37 decreased after treatment with citron essential oils in normal human epidermal keratinocytes (NHEKs); real-time polymerase chain reaction. (B) Protein levels of VEGF and TRPV1 induced by IL-33 and LL-37 in NHEKs decreased after treatment with citron essential oils; enzyme-linked immunosorbent assay. Bars indicate standard deviations. GAPDH: glyceraldehyde 3-phosphate dehydrogenase. *p<0.05, **p<0.01.
![]()
DISCUSSION
Citron is popular for its health benefits. Recent studies suggest that these various pharmacological effects that inhibit disease are known as antioxidant vitamin C and E, polyphenols and flavonoids that are found in many fruits2526. Naringin, a flavonoid present in many citrus fruits including citron, is non-toxic to humans, has a cholesterol-lowering effect, transforms leukemia cells into normal cells, inhibits the proliferation of breast cancer cells, which are known as anti-inflammatory and natural antimicrobial agents2728. Another flavonoid present in citron, hesperidin, is also known to have beneficial effects, such as antioxidant, hypotensive, and antiallergic effects. It is present in a large amount in citrus peels and inhibits an increase in blood cholesterol concentration29. In addition, citron is rich in limonoids such as limonin and normalin, which are excellent antioxidants3031. Limonin is one of the main components that lend bitterness to citrus fruits32. It has been reported to have antibacterial, antiviral, antinociceptive, anti-inflammatory, and anticarcinogenic effects333435. Further, although citron seeds account for 14% to 16% of the total citron fresh weight and most citron seeds are collected and discarded, they are richer in limonoids than is the whole fruit303136. In our previous experiments, we analyzed the effect of citron essential oils and five other essential oils (lavender, rosemary, remongrass, chamomile, and peppermint oil) on NHEKs activated by VD3. RT-PCR and real-time PCR showed that citron essential oils were effective in inhibiting LL-37 and KLK5 (Supplementary Fig. 3). Our additional analyses showed that citron essential oils are abundant in polyphenol (0.5 mg gallic acid equivalents [GAE]/kg in citron seed essential oil and 3.8 mg GAE/kg in unripe citron essential oil). This finding was consistent with those reported previously. Therefore, in this study, we used citron seed and unripe citron essential oils.
This study demonstrates that citron essential oils have a suppressive effect on LL-37, KLK5, TRPV1, and VEGF, which are the main components involved in the pathophysiology of rosacea, in vitro. Augmentation of the cathelicidin innate immune pathway (specifically KLK5 and LL-37) has been reported to be a major contributor to the pathophysiology of rosacea. Rosacea patients have an increased baseline expression of KLK5 and LL-37. KLK5 is the major serine protease responsible for cleaving cathelicidin into its active form, LL-37910. In rosacea-affected skin, LL-37 is processed into shorter fragments that regulate processes such as leukocyte chemotaxis, angiogenesis, and expression of extracellular matrix components9113738. Therefore, agents suppressing the production of KLK5 and thereby inhibiting LL-37 could markedly affect inflammation associated with rosacea. In this study, citron essential oils suppressed the levels of LL37 and KLK5 induced by VD3 in NHEKs. These results support that citron essential oils may normalize the dysregulation of the innate immune system in the skin of rosacea patients and help prevent and improve the symptoms of rosacea.
Recently, TRPV1 and TRP ankyrin receptor (TRPA1) have been found to contribute to the pathophysiology of rosacea. TRPV1 is expressed by sensory nerves and other non-neural cells, such as keratinocytes, and plays a role in vasoregulation and nociception1339. Although the signaling pathways of these receptors are not completely understood, they are activated by rosacea trigger factors such as heat, ethanol, and spicy food40. These receptors release important neuropeptides in neurogenic inflammation, such as substance P and calcitonin gene-related peptide41. In patients with rosacea, these receptors are upregulated, resulting in neurogenic dysregulation leading to rosacea symptoms such as persistent erythema and inflammation838. Although the role of angiogenesis in rosacea is controversial, some studies the support role of angiogenesis in rosacea pathophysiology617. Increased VEGF levels have been reported in lesional skin or rosacea, which result in enhanced inflammation and vascular response seen in rosacea1617. In this study, the levels of VEGF and TRPV1 induced by IL-33 in NHEKs decreased after treatment with citron essential oils. These results demonstrate that citron essential oils may improve rosacea symptoms such as flushing and telangiectasia by suppressing neurogenic dysregulation and angiogenesis in the skin of rosacea patients.
ELISA and immunocytofluorescence revealed that citron seed essential oil had a better inhibitory effect on KLK5 induced by VD3 than did unripe citron essential oil. Furthermore, RT-PCR showed that the inhibitory effects of citron seed essential oil on VEGF and TRPV1 were greater than those of unripe citron essential oil, but the difference was minimal and a dose-dependent pattern was not observed. Thus, the two oils showed comparable results in our experiments. Unlike unripe citron, the citron seed is abandoned in the manufacturing processes. Therefore, it is expected that citron seed will be useful economically and environmentally for development of therapeutic agents for rosacea.
There are several limitations to this study. First, no analysis was performed to determine which components of citron essential oils inhibited the rosacea-related factors. In addition, the mechanisms underlying the inhibitory actions remain to be elucidated, and further experiments are required. To address these limitations, an analysis of the ingredients of citron essential oils and additional experiments are underway.
In this study, it was demonstrated that citron seed and unripe citron essential oils suppressed LL-37, KLK5, VEGF, and TRPV1 in NHEKs stimulated with VD3 and IL-33. LL-37, KLK5, VEGF, and TRPV1 are mediators that play important roles in the pathophysiology of rosacea; hence, citron essential oils are expected to improve rosacea symptoms by inhibiting the underlying pathomechanism. On the basis of our results, we anticipate that citron essential oils could be valuable ingredients for an adjuvant therapeutic agent for rosacea.
 XML Download
XML Download