

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Dopamine (DA) neurons in the substantia nigra pars compacta (SNc) as a pacemaker neuron generate action potential regularly. However, various synaptic and neuromodulatory events can affect them, generating diverse firing patterns including firing pauses and burst discharges [12]. Firing rate and patterns are critical in various DA actions related to many brain functions, including motor control, reward processing, reinforcement learning, and drug addiction [34]. Many excitatory and inhibitory neurotransmitters can influence firing activities of DA neurons. Gamma-aminobutyric acid (GABA), the most dominant inhibitory neurotransmitter, often suppresses spontaneous firing of DA neurons completely [567]. It is well known that striatum, globus pallidus, and substantia nigra pars reticulata project GABAergic afferents to SNc, and that almost 70% of these afferents to the SNc DA neurons are GABAergic [89]. Thus, GABAergic synapses might dominantly govern DA neuron activities. Disinhibition by removing background GABAA conductance can evoke burst firing in certain conditions [10] and in a DArgic model neuron [11]. On the other hand, it has been also reported that GABAB receptors are important for generating burst firings in DA neurons in vivo [12].
GABA activates two different types of receptors: GABAA and GABAB receptors. GABAA receptors are fast-acting ligand-gated chloride (Cl−) channels [1314], whereas GABAB receptors are slow-acting G-protein coupled receptors [1516]. Activation of ionotropic GABAA receptors hyperpolarizes membrane potential and inhibits firing activities of DA neurons [171819]. Metabotropic GABAB receptors can activate many downstream targets, including inwardly rectifying K+ channels, voltage-sensitive Ca2+ channels, and adenylyl cyclase [20]. Activation of K+ channels can generate slow inhibitory postsynaptic potentials (IPSPs) and reduce membrane excitability [21]. They can suppress postsynaptic responses [22] and limit action potential backpropagation [232425]. Local blockade of GABAB receptors in SNc DA neurons in vivo has shown ambiguous effects on firing rate, although it can increase firing regularity [726]. However, exact roles of GABA in nigral DA neurons are far from clear. There are many contradictory reports. For example, inhibitory responses of nigral DA neurons by in vivo stimulation of afferents from the striatum, globus pallidus, and substantia nigra pars reticulata are predominantly mediated by GABAA receptors, but nigral DA neurons in vitro express both functional GABAA and GABAB receptors [71226272829]. Therefore, it has been speculated that postsynaptic GABA induced by neural activity changes is dominantly mediated by GABAA receptors, while slow inhibition via GABAB receptor at presynaptic or perisynaptic sites might be involved when synaptic GABA overflows [71516303132]. However, in highly polarized DA neurons, little is known about where and how GABA receptors exactly regulate spontaneous firing in DA neurons.
Acutely isolated SNc DA neurons can be divided into the soma, proximal dendrite, and distal dendrite. They exhibit regular spontaneous firings [233]. By taking this advantage together with employment of local GABA-uncaging, we here report that GABA signals in the soma and dendrites can differently regulate spontaneous firing in SNc DA neurons of the rat. In the soma, GABA suppresses spontaneous firing equally through GABAA and GABAB receptors. However, in the proximal and distal dendrites, GABA suppresses firing in a GABAB receptor-dominant way.
METHODS
Isolation of DA neurons
Sprague-Dawley rats at 9 to 12 days old were subjected to decapitation. Their brains were quickly excised and placed in ‘high glucose solution’ containing 135 mM NaCl, 5 mM KCl, 10 mM HERPES, 1 mM CaCl2, 1 mM MgCl2 and 25 mM D-glucose. The pH was adjusted to 7.3 with NaOH. Midbrain coronal slices of 400 µm in thickness, containing SNc, were obtained using a vibratome (Series 100, St. Louis, MO, USA). Subsequently, SNc regions of slices demarcated by dark color were dissected out with a scalpel blade and placed in fully oxygenated HEPES-buffered saline containing papain (8 U/ml, Worthington) and incubated at 34-37℃ for 20-30 min. Next, tissue segments were rinsed with enzyme-free saline and then gently triturated with a graded series of fire polished micro-Pasteur pipette. Gentle agitation using various sizes of Pasteur pipettes produced typical single DA neurons. These isolated cells were then plated onto poly-D-lysinecoated small glass cover slips that were already fitted for a recording chamber.
Immunocytochemistry
Acutely isolated cells on glass coverslips were rinsed twice with phosphate-buffered saline and fixed with 4% paraformaldehyde for 40 min at room temperature. After fixation, these cells were washed with phosphate-buffered saline and then incubated in phosphate-buffered saline containing 2% normal goat serum and 0.1% Triton X-100 for 60 min at room temperature. Cells were then incubated for 2 hours in phosphate-buffered saline containing tyrosine hydroxylase antibodies (diluted 1 : 1000), 2% normal goat serum, and 0.1% Triton X-100. After rinsing three times with phosphate-buffered saline, they were incubated with fluorescence isothiocvanate (FITC)-conjugated goat anti-mouse IgG (Molecular probes) diluted 1 : 1000 in phosphate-buffered saline at room temperature for 1 h. Excess fluorescent antibodies were removed by washing three times with phosphate-buffered saline. Fluorescence images were then obtained using a Zeiss 510 confocal laserscanning microscope (excitation at 488 nm; emission at 505-545 nm). Detailed descriptions of morphologies and staining results have been previously reported [33].
Measurement of electrical activity
A patch clamp recording system (EPC-9, HEKA Electronik, Lambreacht, Germany) was used to measure spontaneous firing activities. Patch pipettes were made from 1.5-mm brocilicate glass (WPI, Sarasota, FL, USA) with a Sutter puller (Model P-97, Sutter Instrument, Novato, CA, USA). The resistance of these patch pipettes was between 2 and 3 MΩ. We employed whole-cell and cell-attached configurations in current-clamp mode. In cell-attached and whole-cell mode patch-clamp experiments, electrical signals were continuously sampled at 2 kHz (1 kHz filter). In cellattached mode recording, patch pipettes were filled with normal bath solution. Frequency calculations of spontaneous firing were performed with Igor version 4. (Igor Wavemetrics, Lake Oswego, OR, USA). Interspike interval (ISI) was calculated by counting spike numbers during 1 min. Some data were analyzed using Origin version 6.0 (Microcal Software, Inc., Northampton, MA, USA). When we measured GABA-mediated currents in wholecell configuration, patch pipettes were filled with an KCl-rich internal solution that consisted of 123 mM KCl, 6 mM NaCl, 1.3 mM CaCl2, 1 mM MgCl2, 10 mM EGTA-2K, 10 mM HEPES, 2 mM Mg-ATP, 10 glucose, adjusted to pH 7.3 with KOH.
Uncaging experiments
To increase local GABA within a small area of a single DA neuron, the local uncaging function of the Zeiss 510 confocal microscope was used. We used UV laser (lines 351 and 364 nm) and a 40× oil immersion objective lens (NA=1.3) (Carl Zeiss AG, Germany) in this experiment. Bath solution contained 20 µM (O)-(CNB-caged)-GABA caged-compounds.
Solution and chemicals
Stock solutions of chemicals were prepared with dimethylsulfoxide or triple-distilled water. They were applied in a normal bath solution at known concentrations immediately before using them. The normal bath solution contained 140 mM NaCl, 5 mM KCl, 10 mM HEPES, 10 mM glucose, 1 mM CaCl2, 1 mM MgCl2. The pH and osmolarity were adjusted with NaOH at room temperature (20-24℃) to 7.35 and about 300 mOsm, respectively. Among chemicals related to ionotropic/metabotropic GABA receptors, 6-amino-3-(4-methoxyphenyl)-1(6H)-pyridazin ebutanoic acid hydrobromide (SR95531 hydrobromide, a GABAA receptor antagonist), (2S)-3-[[(1S)-1-(3,4-dichlorophenyl) ethyl]amino-2-hydroxypropyl] (phenylmethyl) phosphinic acid hydrochloride (CGP55845 hydrochloride, GABAB antagonist), 1,2,3,6-tetrahydro-4-pyridinecarboxylic acid hydrochloride (isoguvacine hydrochloride, a specific GABAA receptor agonist), and (R)-4-amino-3-(4-chlorophenyl)butanoic acid (a selective GABAB receptor agonist) were obtained from Tocris Bioscience. UV photolysis of γ-aminobutyric acid, α-carboxy-2-nitrobenzyl ester, trifluoroacetic acid salt (O-(CNB-Caged) GABA) were obtained from Invitrogen.
RESULTS
GABA-induced inhibition of spontaneous firing in SNc DA neurons
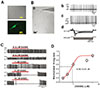
Acutely isolated DA neurons from the SNc show large sized soma attached with 3-6 multiple long dendrites. They exhibit spontaneous firing in normal bath solution [233]. In such large cells, most neurons were immunopositive to tyrosine hydroxylase (TH) (Fig. 1A) and characteristic electrical and morphological features of DA neurons were observed as shown in Fig. 1B. In both on-cell and whole-cell current-clamp modes, regular spontaneous firing activities were recorded without any external stimuli (Figs. 1B-b,c). Step current injections evoked typical ‘sag’ potentials which quickly returned to resting membrane potential (Figs. 1B-d) due to activation of Ih current [34].
To understand how GABA regulates spontaneous firing in this spontaneously firing DA neuron,, we first applied GABA in various concentrations (Figs. 1C and D). Spontaneously firing activities were measured using cell-attached current-clamp condition. GABA at 0.3 µm did not inhibit spontaneous firing. However, at concentration between 0.5 and 1 µM, it suppressed spontaneous firing without a complete blockade of spontaneous firing. At concentration of more than 5 µM, GABA completely blocked spontaneous firing, although there were differences in initial delays depending on GABA concentration applied (Fig. 1C). By measuring numbers of firing during 1 min in several cells, we were able to draw a dose-response curve of GABA-induced firing inhibition. Its IC50 was 0.87 µM (Fig. 1D).
Inward and outward currents evoked by GABAA and GABAB receptors in SNc DA neurons
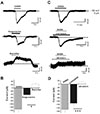
Next, we examined GABA-induced currents after activation of either GABAA or GABAB receptors in acutely isolated DA neurons (Fig. 2). Whole-cell voltage-clamp experiments were carried out using isolated DA neurons. GABAA or GABAB receptor-mediated currents were measured by applying specific GABAA or GABAB receptor agonists to these acutely isolated DA neurons at −50 mV with a KCl rich pipette solution (Fig. 2). As shown in Fig. 2A, GABA evoked an inward current in all cases (−131.6±17.0, n=7). Isoguvacine (20 µM), a specific GABAA receptor agonist, also induced inward currents in all cells tested (−42.4±5.9 pA, n=4). However, (R)-baclofen (100 µM), a specific GABAB receptor agonist, evoked outward currents (14.8±2.8 pA, n=6, Fig. 2A), suggesting the presence of functioning GABAA and GABAB receptors in DA neurons. These results are compatible with previous reports showing that GABAA receptor is a Cl− channel and that GABAB receptors can activate K+ channels in DA neurons [31]. Next, we used specific antagonists for GABA receptor subtypes. When GABA was applied in the presence of either 2 µM CGP55845 (a GABAB receptor antagonist), or 5 µM SR95531 (a GABAA receptor antagonist), we were able to record inward currents after blocking GABAB receptors (Fig. 2C, upper panel) and outward currents by blocking GABAA receptors (Fig. 2C, lower panel), very similar to results of agonist experiments (Fig. 2A). These results are summarized in Figs. 2B and D. Mean current amplitudes of GABA with a GABAB receptor antagonist and GABA with a GABAA receptor antagonist were −112.14±16.22 pA (n=5) and 12.06±2.50 pA (n=5), respectively (Fig. 2D). From these results, it could be concluded that the SNc DA neurons express both functional GABAA and GABAB receptors.
Inward and outward currents evoked by GABAA and GABAB receptors in SNc DA neurons
Next, we examined GABA-induced currents after activation of either GABAA or GABAB receptors in acutely isolated DA neurons (Fig. 2). Whole-cell voltage-clamp experiments were carried out using isolated DA neurons. GABAA or GABAB receptormediated currents were measured by applying specific GABAA or GABAB receptor agonists to these acutely isolated DA neurons at −50 mV with a KCl rich pipette solution (Fig. 2). As shown in Fig. 2A, GABA evoked an inward current in all cases (−131.6±17.0, n=7). Isoguvacine (20 µM), a specific GABAA receptor agonist, also induced inward currents in all cells tested (−42.4±5.9 pA, n=4). However, (R)-baclofen (100 µM), a specific GABAB receptor agonist, evoked outward currents (14.8±2.8 pA, n=6, Fig. 2A), suggesting the presence of functioning GABAA and GABAB receptors in DA neurons. These results are compatible with previous reports showing that GABAA receptor is a Cl− channel and that GABAB receptors can activate K+ channels in DA neurons [31]. Next, we used specific antagonists for GABA receptor subtypes. When GABA was applied in the presence of either 2 µM CGP55845 (a GABAB receptor antagonist), or 5 µM SR95531 (a GABAA receptor antagonist), we were able to record inward currents after blocking GABAB receptors (Fig. 2C, upper panel) and outward currents by blocking GABAA receptors (Fig. 2C, lower panel), very similar to results of agonist experiments (Fig. 2A). These results are summarized in Figs. 2B and D. Mean current amplitudes of GABA with a GABAB receptor antagonist and GABA with a GABAA receptor antagonist were −112.14±16.22 pA (n=5) and 12.06±2.50 pA (n=5), respectively (Fig. 2D). From these results, it could be concluded that the SNc DA neurons express both functional GABAA and GABAB receptors.
Inhibition of spontaneous firing by activation of GABAA and GABAB receptors in DA neurons
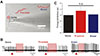
To investigate how effectively GABAA and GABAB receptors could inhibit spontaneous firing activities in SNc DA neurons, specific GABA receptor agonists and antagonists were used, because it was previously confirmed that they could specifically block one type of GABA receptors completely in DA neurons. Bath application of either isoguvacine (20 µM) or (R)-Baclofen (100 µM) inhibited spontaneous firing of DA neurons (Fig. 3A). Their inhibitory effects on spontaneous firing activity were not significantly different (Fig. 3B). The firing rate in control was 2.15±0.68 Hz (n=9). Spontaneous firing rates in the presence of isoguvacine or (R)-baclofen were 0.47±0.18 Hz (n=6) or 0.32±0.17 Hz (n=3), respectively (Fig. 3B). Effects of antagonists for GABA receptor subtypes were also examined. In the presence of CGP55845 (2 µM) or SR95531 (5 µM), GABA was applied for 100 sec (Fig. 3C). Inhibition of one type of GABA receptors, either GABAA or GABAB receptors, did not interfere with GABA-induced inhibition of firing in DA neurons (Fig. 3C). After complete blocking either GABAA or GABAB receptors, GABA dramatically blocked or slowed spontaneous firing rate to 0.28±0.37 Hz (n=3) and 0.41±0.03 Hz (n=2), respectively (Fig. 3C). Results are summarized in Fig. 3D. These results indicate that either GABAA or GABAB receptor alone can inhibit spontaneous firing completely.
Inhibition of spontaneous firing by GABA receptors in the soma and dendritic regions of DA neurons
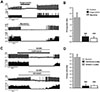
To investigate whether GABA receptors in the soma or dendrites could regulate spontaneous firing differently in polarized DA neurons, we employed local GABA-uncaging techniques in cell-attached recording condition in which intracellular condition was maintained without any violation via dialysis with pipette solution (Fig. 4A). As shown in Fig. 4A, a typical patch-attached DA neuron had long dendrites. To stimulate similar areas of membrane of the soma as much as those in areas of each dendritic region in proximal and distal dendrites, uncaging areas of GABA were adjusted with the same laser expose time (1.50 msec). For this experiment, isolated neurons having long dendrites of >140 µm were selected as shown in Fig. 4A. Bath application of inactivated (O)-CNB caged GABA (20 µM) did not significantly affect the frequency of spontaneous firing (n=3, data not shown). In this typical neuron, local GABA-uncaging sites are seen as red areas in Fig. 4A. Photolysis of caged-GABA on the soma (ISI=2.69 s, a), proximal dendrite (ISI=3.73 s, b), and distal dendrite (ISI=2.33 s, c) induced complete inhibition of spontaneous firing (Fig. 4B). Red triangles in Fig. 4B mean uncaging time while red colored areas indicate duration of firing inhibition. In six neurons, we found that GABA exposures to any area of a DA neuron inhibited spontaneous firing completely. There were no significant differences (p>0.05) in firing inhibition duration among the soma (3.00±0.22 s, n=6), proximal dendrite (3.72±0.70 s, n=6), and distal dendrite (3.10±0.38 s, n=6) (Fig. 4C). These results indicate that GABA can suppress spontaneous firing by GABA receptors not only in the soma, but also in the part of proximal and distal dendrite of the DA neuron.
Different contribution of GABAA and GABAB receptors in the soma and dendrites to firing inhibition in DA neurons
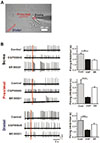
To investigate how GABAA and GABAB receptors in the soma and dendritic tree might participate in the firing inhibition of DA neurons, local GABA uncaging using (O)-CNB caged GABA under the presence of specific blockers for GABAA or GABAB receptors was performed. Red areas in Fig. 5 indicate uncaging sites in the soma, proximal, and distal dendrites. After local GABA uncaging, we repeated the same experiment in the presence of either a specific GABAA receptor blocker or a specific GABAB receptor blocker. In the presence of each antagonist for GABAA and GABAB receptors, GABA uncaging at the soma inhibited spontaneous firing similarly (Fig. 5B, top panel). There was no significant difference in receptor-induced firing inhibitions between GABAA and GABAB (GABAA: 1.97±0.03 s, GABAB: 1.83±0.17 s, n=8, Fig. 5B, top panel), suggesting that GABAA or GABAB receptor alone could inhibit spontaneous firing equally in the soma. In contrast, GABA-induced firing inhibitions in dendritic compartment differed from that of the soma. In proximal dendrites, GABAB receptors (3.02±0.09 s, n=8) inhibited spontaneous firing more strongly than GABAA receptors (1.20±0.06 s, n=8, Fig. 5B, middle panel). Responses at distal dendrites were the same as inhibition of firing at proximal dendritic compartment. ISI was 1.32±0.07 s by GABAA receptor activation (n=8). It was 2.38±0.22 s by GABAB receptor mediated firing inhibition (n=8, Fig. 5B, bottom panel). The bar graph shows grade of firing inhibition (isi) at the soma, proximal dendrite, and distal dendrite (red bars=GABAA antagonist, blue bars=GABAB antagonist) (n=8). These data indicate that in the soma GABAA or GABAB receptors can equally suppress spontaneous firing. However, in the dendrites, GABAB receptors can dominantly suppress spontaneous firing.
DISCUSSION
SNc DA neurons generate spontaneous firing in vivo, in vitro, and under dissociated conditions [235363738]. DA neurons in vivo fire spontaneously in three distinct modes; a tonic regular firing, a random pattern, and a burst firing pattern [35363738]. Because firing modes and/or modulation of regularly firing activity of DA neurons determine DA release, it is very important to understand how excitatory and inhibitory neurotransmitters modulate spontaneous firing activity. Although burst firing is well known to be evoked by glutamatergic afferents [10], it can be also enhanced or evoked by disinhibition of GABAergic afferents in a certain condition [2632]. GABA is a major inhibitory neurotransmitter in the brain. Activation of GABA receptors is known to inhibit neuronal cell activities, but it affects all of the above three firing modes in DA neurons [5639]. GABAergic inputs account for 70% of afferents to the SNc DA neurons [8]. They play a key role in switching of firing patterns in DA neurons [6]. Direct exposure to GABA containing solution inhibits spontaneous firing of DA neurons. This has been reported in brain slices and isolated neurons [1227]. It is also known that nigral DA neurons express both GABAA and GABAB receptors and that application of GABAA or GABAB receptors agonists can inhibit spontaneous firing in vitro [712272829]. However, inhibitory responses of SNc DA neurons by stimulation of afferents from striatum, globus pallidus, or substantia nigra pars reticulata in vivo have been shown to be mediated predominantly by GABAA receptors [32]. GABAergic neurons in the striatum, globus pallidus, and pars reticulata are fast pacemakers that continuously suppress nigral DA neurons. Input activity from the GABA neurons appear to be translated linearly to spike frequency and it is thus likely that this basal ganglia output is frequency coded [151920]. Since both GABAA and GABAB receptors can mediate the inhibitory action of GABA in most neurons including DA neurons [273241], regional distribution of GABAA and GABAB receptors on the soma and/or dendrites in DA neurons appears to be very important for the generation of specific firing patterns and integration of synaptic inputs. In this situation, it is very interesting to see that local electrical stimulation with high frequency trains in vitro evokes both GABAA and GABAB receptors, but that weak stimulation in vivo appears to be mediated exclusively by GABAA receptors [3241]. Therefore, the effect of GABA on spontaneous firing in DA neurons should be determined by strength of GABA afferents as well as location of receptor subtypes. Although GABAB receptors have been primarily studied at the cell body, they are abundant at spines and dendrites [4243] which are known to receive most inhibitory inputs. However, due to the lack of information about distribution of GABAergic buttons and receptors on the somatodendritic tree of DA neurons, it is difficult to exactly estimate how GABA affects firing activities in detail at the moment. In addition, apart from firing regulation, GABA-related mechanisms appear to be involved in neuronal regeneration and axon guidance in the brain [531] as well as in addictive drug actions and reward/aversion pathways [29]. However, GABAA and GABAB type receptor distribution has not been thoroughly tested in DA neurons at single neuron level. Therefore, using patch-clamp recording and local GABA uncaging techniques, we investigated regional roles of GABAA or GABAB receptors in SNc DA neurons.
As a result, in acutely dissociated DA neurons, we confirmed that GABA had inhibitory effect on spontaneous firing. At low concentrations (<5 µM), GABA suppressed tonic firing and reduced spontaneous firing rate. However, at higher concentrations (≥5 µM), it completely abolished spontaneous firing (Fig. 1). Activation of GABAA receptors evoked inward Cl− currents, whereas activation of GABAB receptors seemed to activate outward K+ currents (Fig. 2), similar to results reported in DA neurons [151920]. Although suppressive actions of GABAB receptors in vivo appears to be very weaker than those of GABAA receptors [122732], bath application of GABAA/GABAB receptor agonists and antagonists (in this case together with GABA) showed that spontaneous firing activity could be completely inhibited by activation of only one type of GABAA or GABAB receptor (Fig. 3). Finally, we showed that local application of GABA limited on the soma, proximal dendritic, and distal dendritic region using local caged-GABA uncaging technique, was enough to completely inhibit spontaneous firing (Fig. 4). In case of the soma, activation of GABAA or GABAB receptors equally suppressed spontaneous firing. However, in the proximal and distal dendritic regions, GABAB receptors inhibited spontaneous firing more strongly than GABAA receptors (Fig. 5). This regional difference in the inhibition of GABA receptors between soma and dendritic compartments can help to resolve many complex actions of GABA on spontaneous firing and firing patterns and to understand GABA signaling in DA neurons.
 XML Download
XML Download