

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Endometriosis (EM) is categorized by the growth of functional endometrium tissue exterior to uterus, commonly around the pelvic peritoneum and ovaries, affecting 5–10% of women in their reproductive age [12]. EM is associated with pelvic pain, infertility, dysmenorrhea, and dyspareunia [3]. Present treatment for EM is through surgery and/or hormonal therapy. Hormone treatment involves administration of progestogens, oral contraceptives, and gonadotropin-releasing hormone agonists [4]. However, these therapies are associated with serious adverse effects. Also, lesions reappear in 30–50% women within 3–5 years after surgery [5]. Identification of effective novel additional strategies in the treatment of endometriosis is of immense clinical value.
Understanding the pathogenesis of EM is crucial in development of effective treatment approaches. The development of endometrial lesions involves recruitment of blood vessels via angiogenesis to guarantee supply of oxygen and nutrients [6]. VEGF (vascular endothelial growth factor) is one of the chief pro-angiogenic factor that is involved in endometriosis [7]. VEGF exerts its effects via attaching to its receptors - as VEGF-A and VEGFR-2. Highly proliferative endometriotic lesions exhibit increased expression of VEGFR-2 and VEGF-A in the stroma and in blood vessels respectively [8].
Oxidative stress and inflammatory responses have also been said to be associated with the pathogenesis of EM [910]. Proinflammatory mediators as cytokines and prostaglandins are found to promote EM [111213]. Nuclear factor-kappa B (NF-kB), one of the chief transcription factor of the inflammatory process modulates various processes such as apoptosis, cell proliferation, adhesion, invasion and also angiogenesis in several cell types [14]. These cellular processes are documented to be crucially involved in endometriosis [291415]. Endometriotic lesions expresses increased cyclooxygenase-2 (COX-2) levels than eutopic endometrium [16]. Also, NF-κB induces COX-2 expression in the endometrial stromal cells [171819]. Thus, compounds that inhibit angiogenesis and the COX-2/NF-κB signalling pathways could be potential candidates in the treatment of EM.
Zingiber officinale (Ginger), is widely used in Asian cuisines and also as medicine for various ailments. Gingerols and its derivatives found in the rhizomes of ginger possess numerous biologically active properties [202122]. Shogaols, are gingerols that are found primarily in dried ginger [2023]. 6-shogaol reportedly exhibits wide range of pharmacological activities as antioxidant [24], anticancer [2526], neuroprotective [27] and anti-inflammatory effects [2829]. In this study the effects of 6-shogaol on the major pathways in endometriosis was explored.
METHODS
Experimental animals
Eight to nine weeks old Sprague-Dawley (Female) rats weighing 180–220 g were procured from the laboratory study animal care center of WuHan University and were housed under controlled environmental conditions (12 h light/dark cycle; 22±1℃; 55–60% humidity). Animals had unrestricted access to food and water and were acclimatized to in-house conditions for a week preceding to experiment. Approval of the animal study experiments were obtained from the WuHan University Animal Care and Use Committee and strict NIH Guidelines for the Care and Use of Laboratory Animals were followed [30].
Reagents and antibodies
6-Shogaol (Sigma-Aldrich, St.Louis, MO, USA), Fetal bovine serum (FBS) (Thermoscientific, USA), RPMI1640 medium (Gibco, Grand Island, NY, USA), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT), polyvinylidenedifluoride (PVDF) membranes and enhanced chemiluminescence (ECL) kit (Invitrogen) were used in the study. ELISA kits for determining cytokines-TNF-α, IL-1β and IL-6 were purchased from Biolegend (San Diego, CA, USA). Buffers used in Western blotting analysis were procured from Beyotime Institute of Biotechnology (Beijing, China). Antibodies against VEGF, VEGF-A, VEGFR-2 and COX-2 were procured from Cell Signaling Technology (Danvers, MA, USA). Horseradish peroxidase-labelled IgG secondary antibodies, PGE2, TNF-α, NF-κB p65, IκBα, p-IκBα, p-IKKβ, IKKβ, p-IKKα, IKKα and β-actin were purchased from Santa Cruz Biotechnology (Texas, USA).
Study design
A total of 75 rats (n=15/group) were used in this study. The induction of experimental endometriosis was done as described by Vernon and Wilson [31]. Briefly, a 3 cm midline incision in the abdomen was made to expose the uterus after anesthetizing the rats (i/m of xylazine and ketamine) and one uterine horn was excised and was split longitudinally and sectioned (5×5 mm pieces). The sections were transplanted into the peritoneum and rats were allowed to recuperate. Following 5 weeks after implantation, the spherical volume of ecto-uterine tissues was measured. The animals were randomly grouped into 6 experimental groups. Separate groups of rats were given 6-shogaol (50, 100 or 150 mg/kg b.wt) orally via gavage everyday 24 h following implantation for 30 days. The gestrinone (GTN) group were given GTN (0.5 mg/kg/day) [32] as positive control. Control group animals were not implanted with ectopic tissues nor were treated with GTN or shogaol but, were administered equal volume of saline. Endometriosis control group were implanted with ecotpic tissues but not administered with GTN or shogaol but given saline as in control group.
After 5 weeks, the treated rats were allowed to fast over-night and were sacrificed by cervical dislocation under isoflurane anesthesia. The endometrial tissues were excised with care and the spherical volume was measured as V (mm3)=height×width×length×0.52 mm [33]. The tissues (n=6/group) were then treated in phosphate buffered saline (PBS) and formalin fixed and embedded in paraffin for immunohistochemical and histological analysis.
Measurement of the ectopic tissues
The spherical volume of the ectopic endometrial tissues was measured using Vernier calipe. The difference in the growth and the of the cells following treatment were determined as volume change (mm3)=volume of EMS control (mm3)-Treatment volume (mm3).
Histology and immunohistochemistry
Paraffin-embedded tissues were sectioned into 4 µm thickness and were hematoxylin and eosin (HE) stained and were observed under light microscope (Leica microsystems, Germany) at 200×magnification and analysed.
The expression of VEGF, VEGFR-2 (Flk-1), and COX-2 were assessed by immunohistochemistry. The sections were incubated with specific antibodies overnight (4℃). Following incubation for 40 min with secondary antibodies, the tissues were washed thrice with PBS and were treated with avidin-biotinylated peroxidase complex (ThermoFischer Scientific) for 40 min followed by diaminobenzidine (ThermoFischer Scientific). The tissue sections were then observed and quantified for positive cells with image processing and analysis software (NIS-Elements BR, Nikon Corporation, Japan).
Peritoneal fluid was collected from animals (n=6) after sacrifice by rinsing the abdominal cavity with PBS. The fluid was immediately centrifuged for 10 min (1,500 rpm) and supernatant was collected and stored at −70℃ until use.
Determination of inflammatory mediators by ELISA
Levels of IL-6, IL-1β (Biolegend, San Diego, CA, USA) and PGE2 (Cayman Chemical, Ann Arbor, MI, USA) were determined as per manufacturer's instructions. The levels were analysed using SpectraMax 190 automatic plate reader and were further analysed using SoftMax pro software (Molecular Devices, Sunnyvale CA, USA).
Nitric oxide determination
Nitric oxide levels in the peritoneal fluid was assessed using nitrate/nitrite kit (Cayman chemical, Ann Arbor, MI, USA). Nitrate reductase aids in converting nitrate to nitrite and by addition of Griess reagent nitrite produces a deep purple azo compound. At 540 nm, the compound was measured using ELISA reader. The absorbance of the azo compound accurately reflects nitrite concentration.
Immunoblotting
Using RIPA lysis buffer [50 mM Tris-HCl (pH 7.6), 1% NP-40; sodium deoxycholate (0.5%), SDS (0.1%), PMSF, Aprotinin (1 mg/L), Leupeptin (1 mg/L)] the endometrial tissue sections were homogenized on ice and centrifuged. Total protein concentration in the supernatant was determined using BCA assay kit (BioRad, USA). For NF-κB (p65) expression analysis both in nuclear and cytosolic fractions, aliquot of the tissue homogenate was separated into fractions as per manufacturer's protocol using an NE-PER nuclear and cytoplasmic extraction reagents kit (Pierce Biotechnology, Rockford, IL, USA). Protein concentration of equivalent volumes (50 µg) from each group were electrophoretically separated on SDS-PAGE (10%). The bands were transferred onto PVDF membrane (Invitrogen). The PVDF membrane was then blocked with TBST buffer (20 mM Tris -pH7.6; 137 mM NaCl; 0.1% Tween 20) that contained 5% non-fat milk at 37℃ for 60 min and were incubated overnight with respective primary antibody (1:1,000) at 4℃. The PVDF membranes were washed with TBST thrice and were incubated at 37℃ with secondary HRP-labelled antibodies (1:2,000) for 60 min. Positive bands were analyzed by chemiluminescence method (Millipore, USA) and analyzed by ChemiDoc XRS imaging system (Bio-Rad, USA). Using internal control (β-actin), the concentration of test proteins was normalized.
RT-PCR
RT-PCR analysis was done to explore the influence of 6-shogaol on the levels of VEGF and Flk-1 m-RNA by TaqMan RT-PCR. The endometrial tissue total RNA was extracted as per manufacturer's protocol using TRIZOL reagent (Invitrogen, Carlsbad, CA, USA). Total isolated RNA (2 µg) was used as a template for synthesising cDNA and by using the SuperScript II reverse transcriptase kit (Invitrogen) the first strand was synthesized. With SYBR green fluorescence PCR was carried out using 7300 Real-Time PCR System (Applied Biosystems). The following primers were used for amplification - Flk-1-sense: 5′-GCACTGAATTATGGGAGA-3′, antisense-5′-ATGTGATTTTCTTCTTGATG-3′; VEGF-sense: 5′-ACCATGAACTTTCTGCTC-3′, antisense-5′-GGACGGCTTGAAGATATA-3′; GAPDH-sense: 5′-CACCACCATGGAGAAGGC-3′, antisense: 5′-CCATCCACAGTCTTCTGA-3′. The PCR products were then separated on agarose gel electrophoresis (2%). The bands were stained with ethidium bromide (0.05%) and the intensities were analyzed by Bio-Gel imagery apparatus (Bio-Rad, USA).
Statistical study
Obtained experimental data were statistically analyzed using SPSS software (version 21.0) (SPSS Inc., Chicago, IL). One-way analysis of variance (ANOVA) was performed for comparing multiple groups followed by Duncan's Multiple Range Test (DMRT) with values at p<0.05 as statistically significant.
RESULTS
6-Shogaol treatment inhibits development of ectopic endometrial tissue
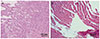
Effect of 6-shogaol on the development of ectopic endometrial tissues were mainly evaluated by the volume change. In the present study volume of the ectopic endometrial tissues of in EMS control group increased significantly (p<0.05) as compared against standard control group. The spherical volume of ectopic tissues was observed to be reduced in rats treated with 6-shogaol (Table 1). GTN exposed animals also displayed decline in the volume as compared against EMS control. Further, the histopathological findings revealed marked atrophy and regressions of endometriotic lesions following 6-shogaol treatment like GTN (Fig. 1).
6-Shogaol effectively inhibits angiogenesis in ectopic endometrial tissues
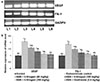
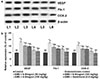
Angiogenesis is well documented as a main process in endometriosis pathogenesis. VEGF and Flk-1 expressions in the endometrial tissues were assessed by immunostaining. The immunological responses were found to be intense in endometriosis control group animals rather in eutopic endometrium of control group animals. Also, mRNA and protein levels of VEGF and Flk-1 as assessed by RT-PCR and western blotting analysis revealed similar observations (Figs. 2 and 3). The results suggest markedly enhanced (p<0.05) angiogenesis process in endometriosis. Further, GTN noticeably decreased VEGF and its receptor Flk-1. It was observed that 6-shogaol treatment for 30 days down-regulated VEGF and Flk-1 expression in a dose-dependent manner indicating inhibition of angiogenesis.
6-Shogaol reduces inflammatory responses
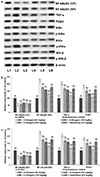
Inflammatory responses are known to be involved in progression of endometriosis. In this study the expression of proteins of the NF-κB pathway, and PGE2 and COX-2 were assessed. Levels of COX-2 expression in immunohistochemical staining and the intensity protein expression as determined by WB (Fig. 4a) were markedly enhanced. NF-κB signalling pathway was noticed to be significantly activated as reflected by elevated expression of NF-κB (p65) in nuclear fraction with considerable (p<0.05) decline in NF-κB (p65) cytosol fraction. Also, up-regulated expressions of PGE2, TNF-α, p-IKKα, p-IκBα and p-IKKβ were noticed (Figs. 4b and c) in endometriotic lesions. Administration of 6-shogaol substantially down-regulated NF-κB (p65) levels in nuclear fraction with substantial (p<0.05) decline in the levels of phosphorylated forms of major kinases of the pathway-IKKα, IKKβ, and IκBα indicating inhibition of the NF-κB pathway. The expressions of PGE2 and TNF-α were also noticeably suppressed on shogaol treatment. Shogaol at 150 mg/kg dose was found to be more effective in down-regulating expressions as compared to lower doses of 50 and 100 mg.
In line with protein expression, the levels of PGE2, IL-1β, IL-6, NO and inflammatory mediators were strikingly raised in the peritoneal fluid of endometriosis induced animals as compared with normal control group (Figs. 5a–c). GTN treatment bring about decline in the levels of PGE2, IL-1β and IL-6. 6-shogaol reduced the levels in a dose-dependent manner. The observations indicate significant ant-inflammatory potential of 6-shogaol.
DISCUSSION
Animal models of endometriosis are extensively employed for understanding the pathology underlying the progression of the disease and for evaluation of effective novel compounds for therapy. The ectopic auto-transplantation of uterine tissues is widely recognized method for inducing endometriosis in study rats [3134]. In this present work, effects of administration of 6-shogaol on growth of implanted endometriotic tissues and on the major pathways of inflammatory process in rats following induction of endometriosis was assessed. 6-Shogaol treatment for 30 days had resulted in declined growth of the endometriotic tissue implants as revealed by the decrease in the volume of the lesions and the histopathological findings indicated significant decrease in atrophy and lesion regression. The observations suggest the efficacy of 6-shogaol.
Inflammatory processes have been crucially involved in endometriosis. Constitutive activation of NF-κB signalling has been noticed in endometriotic lesions and in pelvic macrophages [3536]. NF-κB is found to be a proinflammatory, angiogenic, and a tissue-remodelling factor in endometrial stromal cell (ESC). Activation of the NF-kB pathway was observed in ESC cultures [3738]. COX-2 and MMPs are documented to be up-regulated by NF-κB [3940]. Studies illustrated an NF-kB-dependent expression of inflammatory mediators-COX-2, IL-8, and macrophagemigration inhibitory factor (MIF) in TNF-and IL-1β stimulated ESCs [41424344]. Under normal conditions NF-κB exists in an inactive state found bound to inhibitor proteins (IκB family of proteins) in the cytosol. Upon activation, NF-κBp65 migrates to the nucleus, eventually leading to the expression and release of several pro-inflammatory cytokines [454647]. Peritoneal macrophages exhibiting NF-kB activation has been reported in women with endometriosis [1936].
Expression of angiogenic factors including-COX-2, IL-1β, IL-6, TNF-α, iNOS, pro-inflammatory cytokines and VEGF secreted by activated peritoneal macrophages is known to be critically involved in pathogenesis of endometriosis are regulated by NF-κB [48495051525354]. In this study, activation of NF-κB signalling following induction of endometriosis was observed by increased levels of p65NF-κB in the nuclear fraction and increased phosphorylation of the major regulatory kinases of the pathway. The observed increase in levels of IL-6, IL-1β, COX-2, PGE2 and TNF-α following induction of endometriosis illustrate significant NF-κB activation and elevated inflammation. NF-κB mediated release of inflammatory cytokines consequently promote cell survival and induce the more inflammatory reactions in the lesions [19].
Many of the pro-inflammatory cytokines released by activated macrophages exert angiogenic activities [55]. VEGF is regarded to be a major pro-angiogenic factor in endometriosis [7]. In the study significantly, increased expression of Flk-1 and VEGF were observed in endometriotic lesions. Previous studies have also reported raised VEGF levels in the lesions and peritoneal fluid in endometriosis [56]. Increased Flk-1 expressions were reported by Wang et al. [57]. 6-Shogaol distinctly down-regulated the VEGF and Flk-1 expression both at mRNA and protein levels illustrating inhibition of angiogenesis. This reduced expression could have indirectly aided in reduction in viability of the ectoendometriotic tissues. Further, IL-1β, IL-6 and TNF-α induce VEGF expressions [3454]. Celik et al. [58] reported a significant reduction in microvessel density in a rodent endometriosis model by hindering the NF-κB pathway.
Here, raised levels of cytokines would have in part resulted in elevated VEGF levels and increased cell growth and survival. Studies have demonstrated that the decrease in IL-1β, down-regulates the COX-2 levels in ectopic endometria and inhibits PGE2 expressions that eventually can result in the decrease expressions of estrogen [59]. Also, PGE2 has been reported to induce VEGF expressions thereby contributing to angiogenesis in endometriosis [116061]. The results observed were in line with earlier reports [345462] suggesting the significance of the microenvironment of inflammatory mediators and angiogenic factors in progression of endometriosis. Inhibition of inflammatory responses via down-regulating NF-κB signalling and angiogenesis are critical in therapy of endometriosis.
Several studies have suggested that reduction in proliferation of endometriotic lesions involves the use of hormone therapies, anti-inflammatory drugs as COX-2 inhibitors and TNF-α inhibitors, inhibitors of MMPs, and anti-angiogenic agents [636465]. Here in the present study, 6-shogaol treatment significantly reduced levels of cytokines, PGE2, VEGF, COX-2 and NO that strikingly demonstrates the anti-inflammatory effects of shogaol. The results illustrate efficacy of 6-shogaol against progression of endometriosis via down-regulating growth, VEGF-mediated angiogenesis and NF-κB-mediated inflammation. Ergenoğlu et al. [66] demonstrated reduction in implant size and histological changes in the endometrial lesions following treatment with resveratrol. Administration of curcumin was found to decrease the ectopic endometrial glands [67]. These reports indicate the use of phytomedicines in endometriosis and, also support the results of the present investigation, thus making 6-shogaol a potential candidate drug that could be investigated more in therapy of endometriosis.


 XML Download
XML Download