

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
It is well known that, in adults, the pulmonary pleura forms an evenly smooth lung surface. The pleura transmits the respiratory forces to the underlying lung parenchyma without frictional damage [1]. From the third fetal trimester onward, the lung surface has been shown to develop an evenly smooth morphology to adapt to respiratory movements in utero to facilitate a further development of the airway and diaphragm [2]. However, during pseudoglandular and canalicular phases in midterm fetuses, the lung surface might be shaped differently due to proliferation, branching and outward growth of the bronchiolar system. At its costal surface, the fetal lung lies adjacent to the thoracic wall, while its basal surface is located adjacent to the diaphragmatic dome and shifted cranially by the growing liver. Thus, it is likely that the shape of the lung in fetuses, at least at its costal and basal aspects, is determined by the shape of the thoracic cavity, which is firm enough to resist the pressure of the growing and expanding lungs. Hayashi et al. [3] described that in human embryos and fetuses at 6–7 weeks of gestation, a sheet of mesenchymal tissue that lines the internal aspects of the thoracic cavity gradually disappears to increase capacity of the pleural cavity. Therefore, until 8 weeks, the growing lungs seem to determine the shape of the early pleural cavity, to adjust it to the lung surface. However, there is no systematic description as to when and how the costal and basal surfaces of the fetal lung become smooth.
Here, we conducted a systematic analysis of the development of the pulmonary pleura of human fetuses at 9–16 and 26–34 weeks of gestation. Unfortunately, because of long-lasting preservation in formalin solution, the present specimens were not suitable for immunohistochemistry. However, according to Abe et al. [4] and Katori et al. [5], CD34 antibodies were likely to be useful for identification of fetal fibrous tissue configurations. Although CD34 is widely known as a marker of vascular endothelial cells and hematopoietic stem cells, it is well suitable to identify fibrocytes in specific locations such as skin and Tunica externa of lymphatics [67]. In addition, using elastica-Masson staining (a variation of Masson-Goldner staining), we examined whether or not the fetal pleura contains elastic fibers. According to Kinoshita et al. [8], who demonstrated initial appearance of elastic fibers in the fetal head and neck, we expected to find elastic fibers in the lung after gestational week 20. Consequently, the aim of this study was to describe the fetal pulmonary pleura in relation to the lung surface morphology.
Materials and Methods
This study was performed in accordance with the principles of the Declaration of Helsinki 1995 (as revised in 2013). For a better understanding of lobar and segmental topographical anatomy of the lung, we usually chose sagittal from 4 collections of paraffin-embedded histological sections of fetuses at midterm and late stage. We observed sections from 41 fetuses: (1) eleven Chinese specimens at 9–16 weeks (crown-rump length [CRL], 39–125 mm), (2) eight Japanese specimens at 26–34 weeks (CRL, 210–290 mm), (3) seventeen Spanish specimens at 10–15 weeks (CRL, 47–117 mm), and (4) five German specimens at 9–11 weeks (CRL, 39–64 mm). Spanish and German specimens were from serial sections, while those from China and Japan were semi-serial. The sectional plane was sagittal in 30 of 41 specimens, while the other 11 specimens were horizontally sectioned.
Eleven Chinese specimens were donated by the fetuses' families to the Department of Anatomy, Yanbian University Medical College, Yanji, China, until 2016 and the use for research was approved by the university ethics committee in Yanji (No. BS-13-35). These fetuses were obtained by induced abortion, after which the mother was orally informed by an obstetrician at the college teaching hospital of the possibility of donating the fetus for research. No attempt was made to encourage the donation actively. After the mother agreed, the fetus was assigned a specimen number and stored in 10% w/w neutral formalin solution for more than 1 month. Because of specimen number randomization, there was no possibility of contacting the family at a later date. The trunk samples were decalcified by incubating at 4℃ in a 0.5-mol/l EDTA (pH 7.5) decalcifying solution (Decalcifying Solution B, Wako, Tokyo, Japan) for 3–5 days, depending on the size of the sample. After routine procedures for paraffin embedded histology, the specimens were sectioned sagittally (5 specimens) or horizontally (6) at 20–50 micron intervals depending on sizes of the fetuses (thickness of sections, 5 µm).
Eight Japanese specimens were parts of a collection in the Department of Anatomy, Akita University, Akita, Japan. They were donated by their families to the Department during 1975–1985 and preserved in 10% w/w neutral formalin solution for more than 30 years. The available data was limited and included date of donation and gestational week, but not family name, name of obstetrician or hospital, and reason for abortion. The use for research was approved by the university ethics committee in Akita (No. 1428). After removal of lungs, heart and thymus en bloc, we prepared eight paraffin blocks. From each of the blocks, we performed semiserial sagittal sections of 5 µm at 50–200-micron interval, depending on the size of fetuses.
Most of sections from the Chinese and Japanese specimens were stained with hematoxylin and eosin (H&E), while some of them were used for immunohistochemistry as well as Elastica-Masson staining, i.e., a variation of Masson-Goldner staining [910]. The primary antibodies were (1) mouse monoclonal anti-human CD34 (dilution 1:100, IR632, Dako, Glostrup, Denmark); (2) monoclonal anti-human podoplanin (1:100 dilution, Nichirei D2-40, Tokyo, Japan). For the latter, we needed autoclave treatment (105℃, 10 minutes) after immersion in a ligand activator (Histofine SAB-PO Kit, Nichirei). The secondary antibody (incubation for 30 minutes, dilution 1:1,000) was horseradish peroxidase–conjugated anti-mouse IgG (Histofine Simple Stain Max-PO, Nichirei). Enzyme reaction was visualized with 3,3′-diaminobenzidine (3–5 minutes, Histofine Simple Stain DAB, Nichirei). Samples were counterstained with hematoxylin. Negative controls consisted of samples without primary antibody.
Seventeen specimens were part of the large collection at the Embryology Institute of the Universidad Complutense, Madrid, obtained from miscarriages and ectopic pregnancies at the Department of Obstetrics of the University. Most sections were stained with H&E, some with azan, orange G or silver staining. The study was approved by Universidad Complutense ethics committee (B08/374). Finally, five specimens were from the Blechschmidt collection at the Medical Museum of Georg-August-Universität Göttingen, and sectioned sagittally. Sections were stained with HE, azan or Masson trichrome staining. The use of the collection did not require a specific approval of the Institute.
Results
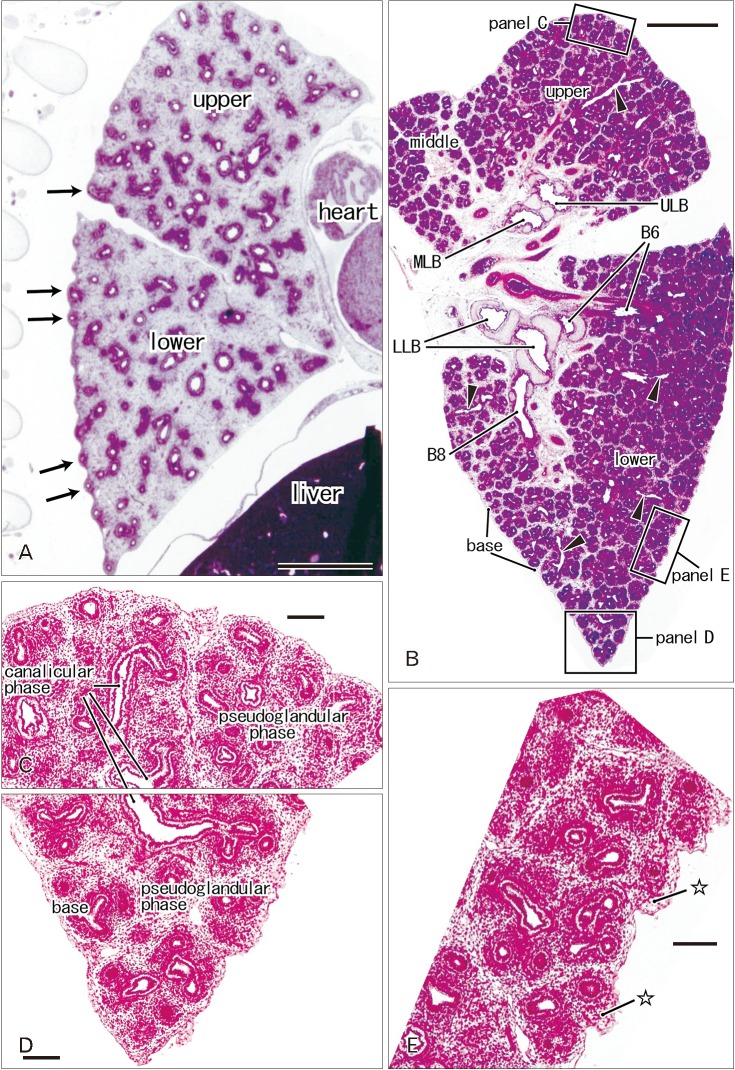
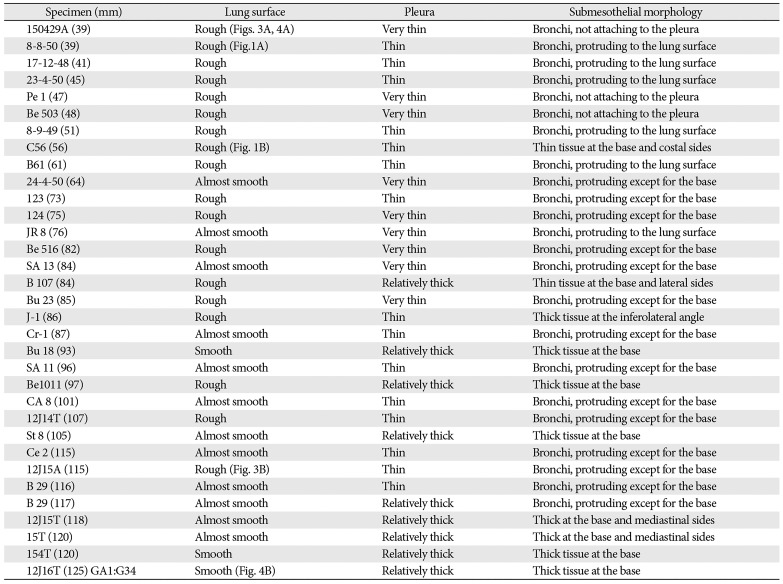
At 9–16 weeks (CRL, 39–125 mm), the pseudoglandular and canalicular phases of lung development were present in various proportions (Fig. 1). The lateral or costal surface as well as the basal surface was usually rough since the outward growing bronchioli provided abundant small protrusions at the surface (Fig. 1A, B). However, whether it was smooth or rough depended on the specimens (Fig. 1A [CRL, 39 mm]). The pulmonary pleura was thin and even difficult to identify (Table 1). Beneath the pulmonary mesothelium, no or little mesenchymal tissue was present at the superficial side of the bronchioli along the costal surface, whereas the submesothelial tissue was relatively thick at basal and medial sites (Fig. 1C–E). No intrapulmonary vein reached the pleural tissue.
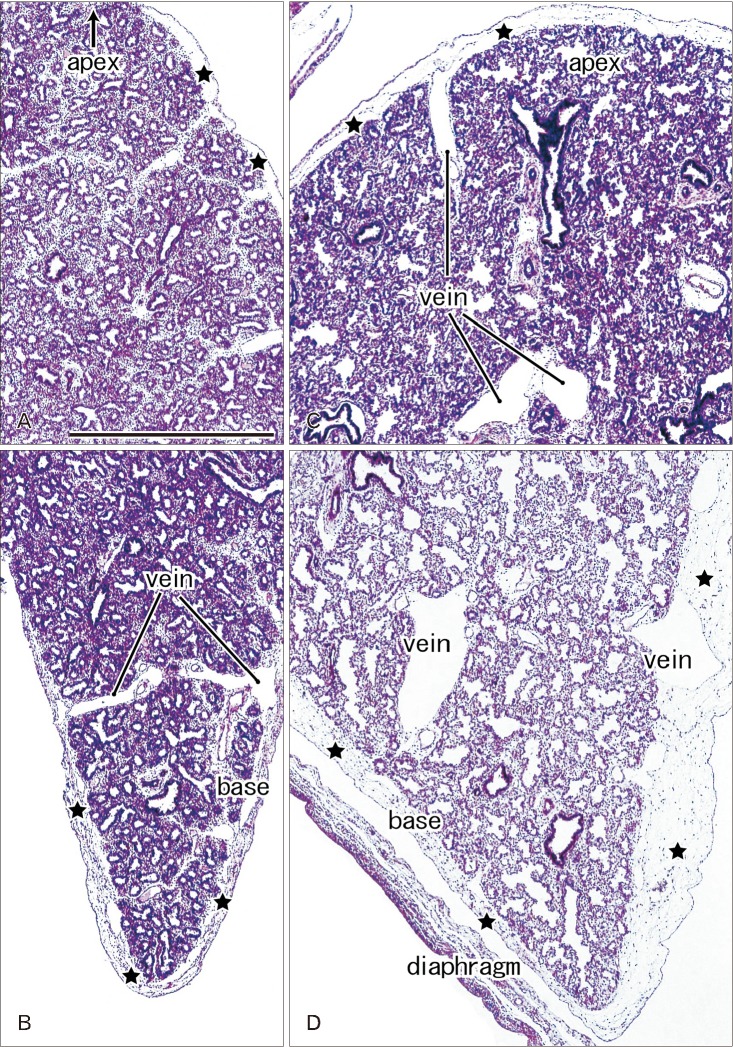
In later stages (fetuses at 26–34 weeks; CRL, 210–290 mm), the lungs were in the saccular or alveolar phases of development and carried almost smooth surfaces because, instead of outward growing bronchioli, the developing alveoli faced the lung surfaces and made them soft and flat (Fig. 2). Moreover, the prominent submesothelial tissue (lamina propria) contained numerous dilated veins connected to the deep intersegmental veins. The deep veins were also dilated. Multilayered fibers were also evident in the lamina propria. The lamina propria was usually thick at the basal and mediastinal areas (Table 1), and sometimes provided a thick fold-like protrusion at the inferolateral angle of the lower lobe.
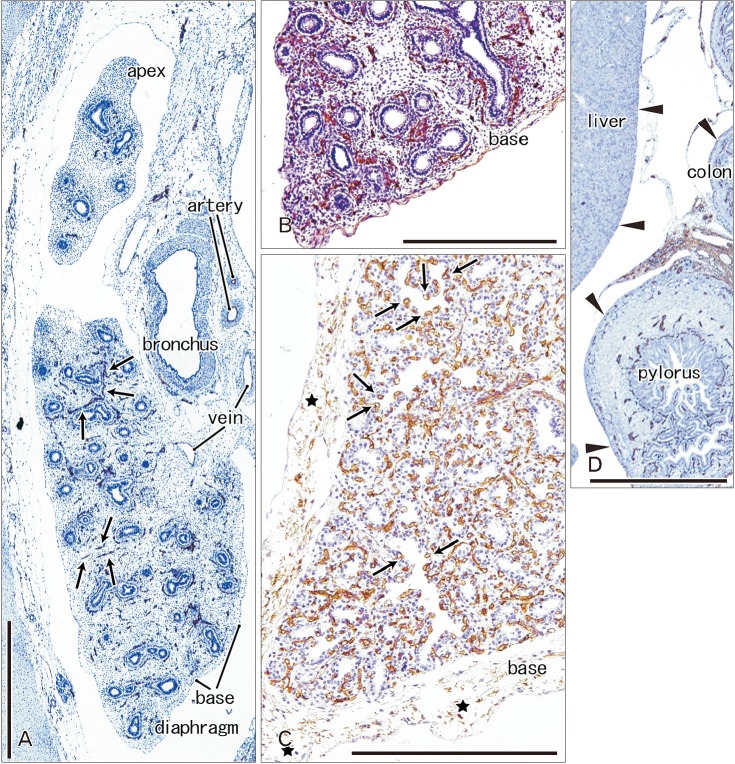
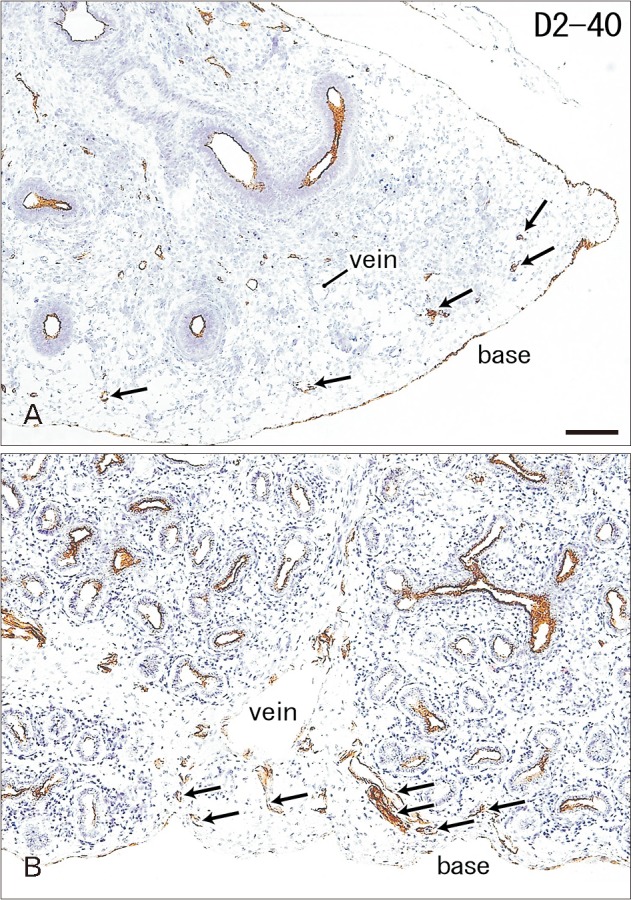
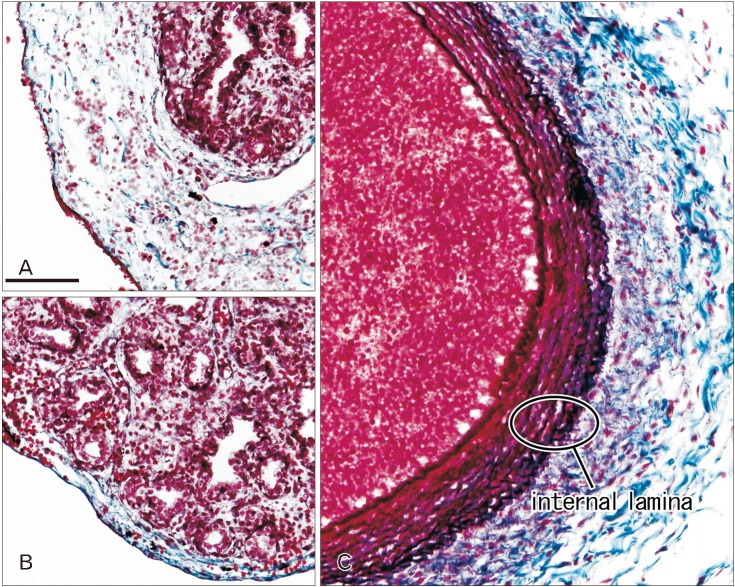
Immunoreactivity for CD34 first appeared in developing vessels between bronchioli (Fig. 3A) and extended to the submesothelial tissue (Fig. 3B, C). Capillaries adjacent to the developing alveoli, characteristic of the canalicular and saccular phases, also expressed CD34 in the late stage fetuses (Fig. 3C). Actually, the subpleural tissue was still thin at 15 weeks (Fig. 3B), but serosal surfaces of the liver and abdominal gastrointestinal tract carried no or little CD34-positive fibers or tissues (Fig. 3D). The pulmonary pleura was weakly positive or negative for CD34: this was same as in the peritoneum. D2-40–positive lymphatic vessels were identified at and near the lung surface even in the smallest specimen (CRL 39 mm) in this study (Fig. 4A). At 16 weeks, the lymphatic vessels were thick and increased in number in the intersegmental fibrous tissue as well as in the submesothelial tissue (Fig. 4B). D2-40 reactivity was not seen in the larger specimens possibly due to long preservation in formalin solution. Finally, we found elastic fibers in larger specimens especially in the internal lamina of any thick arteries (Fig. 5), but the lung contained no or only few elastic fibers. The pleura was stained darkly gray with the elastic fiber staining (Fig. 5A, B), but according to the morphology, the positive structure was different from wavy elastic fibers seen in the arterial wall.
Discussion
In the present study, we demonstrated that smooth lung surface was established until 26 weeks of human fetal development. Instead of the outward growing bronchioli at the pseudo-glandular and canalicular phases, the smooth surface was characterized by increasing numbers of alveoli facing the lung surface. Subsequent development of the submesothelial tissue appeared to make the smooth surface complete. Thickening of the submesothelial tissue seemed to depend on the development of superficial veins that were connected with deep intersegmental veins. Lymphatic vessels developed much earlier than superficial veins, but their volumes seemed to be limited in the submesothelial tissue. The submesothelial tissue is known as a major lymphatic drainage route from the deep, subsegmental parenchyma to the hilar nodes, as a morphological basis for early metastasis of lung cancer [1112].
Actually, the lamina propria of the pleura was often thick at the basal and mediastinal surfaces. However, rather than a possible mechanical stress from the thoracic wall and diaphragm during respiratory movement in utero, the submesothelial tissue including vessels seemed to much contribute to provide the smooth lung surface. Even in full-term babies, the number of alveoli is only 8% of that in adults [13]. Therefore, it is unlikely that the fetal lung occupies the full size of the potential pleural space. A thick fold of the submesothelial tissue at the inferolateral angle of the lower lobe may occupy an excess space of the pleural cavity.
Matrices in the fetal pulmonary interstitium have been of major interest for embryologists with a focus on epithelial-mesenchymal interactions and epithelio-mesenchymal transitions [14151617]. Some components of the extracellular matrix (ECM), such as tenascin-C and others, are likely to mechanistically support branching and growth of bronchioli [17]. Likewise, the pulmonary pleura and submesothelial tissue are likely to contain ECM components against the mechanical contact with the thoracic wall. We were not able to perform immunohistochemistry of ECM components because of a limitation of materials, but using immunohistochemistry of CD34, we demonstrated a fiber configuration of the tissues. CD34-positive fibrous tissues are evident in and along viscera but not in and around skeletal muscles: this fact makes a clear site-dependent difference in distribution [45]. Submesothelial fibrous components were fewer in and along the liver and abdominal gastrointestinal tract than the lung in a same section. The thin submesothelial tissue in the abdomen might be a result of no or few veins and lymphatics contained [1819]. The subpleural fibers might conduct mechanical stress from the thoracic wall to the lung ECM with or without modification.
 XML Download
XML Download