

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Inflammation is the sum of the complex biological responses caused by an overactivated immune system. Inflammation is closely associated with the pathology of many diseases, such as autoimmune diseases like multiple sclerosis and rheumatoid arthritis as well as degenerative diseases like Alzheimer's disease, cardiovascular disease, and cancer.1 Consequently, it will be important to develop treatments that can alleviate or cure these diseases.
Dendritic cells (DCs) perform essential roles in innate and adaptive immunity, such as processing and presenting xenobiotics intruded into the host and acting as a central regulator of the immune responses.2 These functions of DCs are controlled by various pro-inflammatory cytokines, such as interleukin (IL)-12p40, IL-6, and tumor necrosis factor (TNF)-α.3
IL-12 has two subtypes, p35 and p40, which have disulfide bonds in different locations. It is produced by antigen-presenting cells, like DCs, monocytes, and macrophages, and induces an inflammatory response to xenobiotics and immune signals.4 It might also be a trigger that initiates cellular immunity. IL-6 was originally known as a B-cell differentiation factor and is involved in various biological actions, such as the acute immune response and hematopoiesis.5 TNF-α is mainly released by monocytes and macrophages and is a pro-inflammatory cytokine involved in host defenses against xenobiotics and cancer.6 TNF-α might also be involved in the pathogenic mechanisms of acute and chronic inflammatory disease.7
Pro-inflammatory cytokines play important roles in host immunity, but their overexpression can make a disease worse, triggering fever and tissue damage and even leading to death. Suppressing the overproduction of these cytokines could be a way to treat diseases related to inflammation.
Cimicifuga dahurica (Turcz.) Maxim. belongs to the Ranunculaceae and grows widely in Korea, China, and Japan where it is traditionally used as an antipyretic and analgesic.8 This medicinal plant produces cycloartane-type triterpenoids, indolinone alkaloids, phenolics, and their glycosides. Plant preparations are believed to have anti-cancer, anti-inflammatory, and antioxidative effects.9101112 Previously, we reported that components from C. dahurica inhibit soluble epoxide hydrolase, which is one of the causes of cardiovascular and renal disease.131415 As part of our ongoing investigations of the anti-inflammatory activity of Korean medicinal plants, we found that the methanol extract of C. dahurica and dichloromethane and water fractions showed significant in vitro anti-inflammatory activity. Here, we studied its immunoregulatory effect by measuring excreted pro-inflammatory cytokines in the culture supernatant of bone marrow-derived dendritic cells (BMDCs) stimulated by lipopolysaccharide (LPS).
Experimental
Extraction and isolation
The fresh roots of C. dahurica (2.5 kg) were cut into pieces and extracted with 95% aqueous EtOH (3 × 5.0 L) under ultrasonic agitation at 90 Hz and 40℃. The ethanol solution was concentrated under vacuum and filtered through a Buchner funnel to give a brown extract (65.3 g), which was suspended in distilled H2O and successively partitioned with n-hexane and CH2Cl2 to afford n-hexane (9.6 g, A), CH2Cl2 (15.2 g, B) fractions, and a H2O layer (W).
The CH2Cl2 fraction was partitioned by column chromatography over silica gel and eluted with n-hexane-EtOAc (95:5, 80:20, 40:60, v/v) to produce seven fractions (Fractions, B-1 to B-7). Fraction B-3 (0.8 g) was chromatographed by Sephadex LH-20 CC eluted with MeOH-H2O (95:5, 70:30, 50:50, v/v) to give three subfractions (Fractions, B-3.1 to B-3.3) and further purified by YMC RP-C18 CC using acetone-H2O (3:2) as the eluent to afford 24-epi-acerinol (1, 6.8 mg), 23-O-methyl-cimiacerogenin B (2, 3.5 mg), and 25-triepoxy-12β-acetoxy-3β,26-dihydroxy-9,19-cyclolanost-7-ene (3, 10.2 mg). Next, fraction B-4 (1.1 g) was chromatographed over a silica gel CC eluted with n-hexane-EtOAc (2:1) to obtain 7,8-didehydrocimigenol (4, 13.5 mg), 24-epi-24-O-acetyl-7,8-didehydroshengmanol (5, 4.0 mg), and 25-anhydrocimigenol (6, 3.1 mg). Similarly, fraction B-5 (1.3 g) was separated by YMC RP-C18 CC using acetone-H2O (2.5:1) and Sephadex LH-20 using CH2Cl2-MeOH (20:80) as eluent to give 25-O-acetyl-7,8-didehydrocimigenol (7, 5.6 mg). Next, fraction B-7 (1.1 g) was chromatographed on a column of Sephadex LH-20 and eluted with H2O in MeOH, successively, to give three sub-fractions (Fractions, B-7.1 to B-7.3). Subfraction B-7.2 (0.5 g) was separated by CC over silica gel using a mixture solvent of CH2Cl2-MeOH-H2O (8:1:0.08) as eluents and further purified by YMC RP-C18 CC, eluted with MeOH-H2O (4:1), to give ferulic acid methyl ester (832.6 mg).
The H2O fraction was separated using a Diaion HP-20 column and was eluted with a gradient solvent mixture of MeOH-H2O (25:75, 50:50, 65:35, 75:25, to pure MeOH, stepwise) to yield four fractions (W-1 to W-4), based on TLC analysis. When the same steps were repeated as above, 7,8-didehydro-25-anhydrocimigenol-3-O-β-D-xylopyranoside (9, 18.2 mg) was also obtained by purifying subfraction W-2 on YMC RP-C18 silica gel and followed by passage over a Sephadex LH-20 column using mixtures of MeOH-acetone-H2O (8:1:1). Next, fraction W-3 (1.7 g) was subjected to YMC RP-C18 silica gel CC and was eluted with a solvent mixture of H2O-MeOH (5:1, 3:1, 1:1, and 100% MeOH) to afford four smaller subfractions (W-3.1 to W-3.4). Further purification of subfraction W-3.1 (0.42 g) via silica gel column eluted with CH2Cl2-MeOH (7:1), yielded a compound mixture (18.2 mg) that was further purified by preparative TLC (acetone-H2O, 1:2) to yield 24-epi-25-O-acetyl-7,8-didehydrocimigenol 3-O-β-D-xylopyranoside (10, 9.5 mg). In a similar process to that described above, subfraction W-3.3 (0.07 g) was subjected to YMC RP-C18 silica gel CC (MeOH-H2O, 1:1) and then was further purified by silica gel CC using to yield cimiricaside C (11, 10.2 mg). Finally, the fractionation of subfraction W-4 (1.3 g) was separated via silica gel CC and eluted repeatedly with CHCl3-EtOAc (8:1, 4:1, and 2:1) to yield five smaller subfractions (W-4.1 to W-4.5), which were separated by passage over a Sephadex LH-20 column and then applied to repeated silica gel CC (LiChroprep RP-C18 column) and a silica gel CC eluted successively with CH2Cl2-EtOAc-H2O (5:1:0.1) and CHCl3-acetone (1:1) to yield (26R)-cimicifugoside (12, 9.8 mg to yield).
Cell cultures
Bone marrow-derived dendritic cells were grown from wild-type C57BL/6 mice (Orient Bio Inc., Seoul, Korea). All animal procedures were approved by and performed according to the guidelines of the Institutional Animal Care and Use Committee of Jeju National University (#2010-0028). Briefly, the mouse tibia and femur were obtained by flushing with Dulbecco's modified Eagle medium to yield bone marrow cells. The cells were cultured in Rosell Park Memorial Institute (RPMI) 1640 medium containing 10% heat-inactivated fetal bovine serum (FBS; Gibco, Grand Island, NY, USA), 50 µM β-mercaptoethanol, and 2mM glutamine supplemented with 3% J558L hybridoma cell culture supernatant containing granulocyte-macrophage colony-stimulating factor (GM-CSF). The culture medium was replaced with fresh medium every other day. At day six of culture, non-adherent cells and loosely adherent dendritic cell (DC) aggregates were harvested, washed, and resuspended in RPMI 1640 supplemented with 5% FBS.
Cytokine production measurements
The BMDCs were incubated in 48-well plates in 0.5 mL containing 1 × 105 cells per well, and then treated with the extract, fractions and isolated compounds (1 – 12) at various concentrations for 1 h before stimulation with 10 ng/mL LPS from Salmonella minnesota (Alexis, Famingdale, NY, USA). Supernatants were harvested 18 h after stimulation. Concentrations of murine IL-12 p40, IL-6, and TNF-α, in the culture supernatant were determined by ELISA (BD PharMingen, San Diego, CA, USA) according to the manufacturer's instructions. The data are presented as means ± S.D. of at least three independent experiments performed in triplicate. The inhibitory effects are represented as giving 50% inhibition (IC50) relative to the vehicle (DMSO) control. SB203580, an inhibitor of p38 MAP kinase, was used as a positive control.
Result and Discussion
The methanol extract and dichloromethane and water fractions were subjected to cell treatment to evaluate the level of cytokines in LPS-stimulated BMDCs. The methanol extract of C. dahurica inhibited IL-12p40, IL-6, and TNF-α production (IC50 = 2.45, 1.21, and 22.0 µg/mL, respectively; Table 1). Since the methanol extract significantly reduced inflammation, it was partitioned to obtain a dichloromethane-soluble portion and an aqueous phase. As shown in Table 1, the dichloromethane-soluble fraction strongly inhibited LPS-stimulated IL-12p40, IL-6, and TNF-α production (IC50 = 6.82, 0.07, and 11.62 µg/mL, respectively), while the aqueous layer strongly suppressed the production of IL-12p40 (IC50 = 2.43 µg/mL) and TNF-α (IC50 = 2.98 µg/mL) (Table 1).
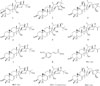
All of the compounds (1 – 12, Fig. 1) isolated from the dichloromethane fraction and water layer of the C. dahurica extract were tested for inhibitory effects on the production of the pro-inflammatory cytokines IL-12p40, IL-6, and TNF-α. The results of 3-(4,5-dimethyl-2,5-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assays showed that compounds 1 – 12 did not exhibit significant cytotoxicity at the concentrations tested (data not shown). The positive control, SB203580, an inhibitor of p38 MAP kinase, inhibited IL-12, IL-6, and TNF-α production with IC50 values of 5.00 ± 0.15, 3.50 ± 0.25, and 7.20 ± 0.21 µM, respectively (Table 2).Of the tested compounds, 1 – 8 and 10 – 12 inhibited IL-12p40 production more strongly than SB203580, with IC50 values of 3.0 µM or less (Table 2). The variability in inhibition of the inflammatory response by 1 – 8 and 10 – 12 may be explained by the different levels of secreted inammatory factors following LPS stimulation. Compound 6 (a cycloartane-type triterpenoid) strongly inhibited LPS-stimulated IL-12p40 production (IC50 = 1.92 µM), at a level comparable to that of SB203580 (IC50 = 5.00 µM), while the inhibitory effect of compound 9 on IL-12p40 production was moderate (IC50 = 16.35 µM). When the sugar unit at C-3 of the aglycone in compound 9 was linked to a xylose chain, the effect was weaker than that of compound 6, suggesting that a hydroxyl group at position C-3 of aglycone is an important functional element.
Regarding IL-6, the methanol extract, dichloromethanesoluble fraction, and water layer strongly inhibited LPS-stimulated IL-6 production (IC50s = 1.21, 0.07, and 24.00 µg/mL, respectively). Of the tested compounds, 3, 4, 7, and 9 – 11 exerted greater inhibitory effects than SB203580 on IL-6 production with IC50 values of 0.45, 0.023, 0.41, 0.01, 0.04, and 1.32 µM, respectively (Table 2). This variability in the inhibition of the inflammatory response by 3, 4, 7, and 9 – 11 may be explained by the secretion of different levels of inflammatory factors upon LPS stimulation.
Overexpression of the pro-inflammatory cytokines TNF-α and IL-6 is associated with the development of autoimmune, inflammatory, and immunopathological diseases. Therefore, blocking TNF-α, IL-6, and their respective signaling pathways might be effective in the treatment of inflammatory diseases. Our results indicated that the water layer inhibited LPS-stimulated TNF-α production (IC50 = 2.98 µg/mL). Notably, compounds 2, 4 – 8, and 12 strongly inhibited TNF-α production (IC50s = 2.71, 3.11, 3.37, 4.60, 4.16, 4.77, and 4.78 µM, respectively; Table 2). Nevertheless, the water layer was more active than these compounds (2, 4 – 8, and 12), suggesting the presence of other active derivatives that acted individually or synergistically with the isolated cycloartane-type derivatives.
The IC50 values of the seven compounds (2 – 5, 7, 8, and 12) that inhibited the pro-inflammatory cytokines IL-12p40, IL-6, and TNF-α in LPS-stimulated BMDCs were much higher. The most promising activity was found with cycloartane-type triterpenoid compounds, which appear to have specific anti-inflammatory activity. It is difficult to recognize clear structure–activity relationships within the series of compounds discovered. However, there was a clear, albeit weak, correlation with their lipophilicity, with higher activity with increasing lipophilicity.
In conclusion, this work is the first to report the inhibitory effect of extracts of C. dahuricaon pro-inflammatory cytokine production by LPS-stimulated BMDCs in vitro. The results support their ethnomedical use in the treatment of inflammation. This report suggests why these medicinal plants possess anti-inflammatory properties and could provide a scientic basis for their use as supplementary herbal products for treating inflammation.


 XML Download
XML Download