

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Innate lymphoid cells (ILCs) are a recently identified group of lymphoid cells that lack an antigen receptor and develop from common lymphoid progenitors in fetal liver and bone marrow.1 ILCs have been classified into 3 groups and each group produces cytokines and transcription factors resembling each of the T helper cell subsets.12 Group 1 ILCs (ILC1s), which consists of classical natural killer (cNK) cells and ILC1s, can produce the Th1 cytokine interferon (IFN)-γ and express T-box transcription factor (T-bet). They have been implicated in Th1-like immunity against intracellular pathogens such as viruses and intracellular bacteria.34 Group 2 ILCs (ILC2s), which consists of natural helper cells and nuocytes (inflammatory ILC2s), can produce Th2 cytokines: interleukin (IL)-4, IL-5, IL-9, and IL-13, and express GATA-3.56789 They have been implicated in immunity against helminths, allergic diseases and tissue repair.1 Group 3 ILCs (ILC3s) consist of the 2 major subsets of natural cytotoxicity receptor (NCR) NCR+ ILC3s and NCR− ILC3s.10 ILC3s can produce the Th17 cytokine IL-17A, but IL-22 can only be produced by NCR+ ILC3s. ILC3s express the retinoid-related orphan receptor γt (RORγt) and play a role in immunity against extracellular bacteria, which are implicated in the pathogenesis of some autoimmune diseases.10
Currently, it has been demonstrated that ILCs, especially ILC2s are involved in allergic airway inflammation. ILC2s in peripheral blood and bronchoalveolar lavage fluid (BALF) were increased in asthmatic patients compared to control subjects and were negatively correlated with airway function.11121314 Many inhalant antigens trigger IL-33 and IL-25 production from epithelial cells and various immune cells, and induce eosinophilic asthma-like airway inflammation through activation of lung ILC2s.1516171819 The strong protease activity of house dust mites (HDMs) induces IL-33 release from epithelial cells, which provokes asthma-like airway inflammation through ILC2 activation.1 In patients with allergic rhinitis (AR), ILC2s are increased in peripheral blood after a cat allergen challenge and during grass pollen season.202122 The levels of ILC2s in HDM AR patients was higher compared to healthy controls and had a positive correlation with plasma IL-13 levels.2324
HDM is globally one of the most common problematic allergens, especially in the tropics.25 In Asia, a recent review reported that the prevalence of HDM sensitization could be as high as 90% or more in some countries such as Thailand, Singapore, Taiwan and India.26 Allergen immunotherapy (AIT) is an important treatment against HDM sensitization in allergic rhinitis and asthmatic patients.2728 However, the effect of AIT on ILCs is controversial.29 AIT can reduce ILC2 levels in HDM AIT patients and grass pollen AIT patients.2130 However, the number of ILC2s were not significantly different before or after 2 or 4 months of AIT between patient undergoing treatment and placebo groups in grass pollen allergy but slightly decreased out of the pollen season.22 No data on circulating ILC1s and ILC3s in HDM-specific immunotherapy study has previously been reported. This study aimed to evaluate the effect of AIT on ILC1s, ILC2s, and ILC3s by comparing them between healthy, AR without AIT, AR with responder and non-responder AIT. We also studied the activation of the ILC2s induced by allergen-specific re-stimulation.
MATERIALS AND METHODS
Study population
The cross-sectional study recruited adult subjects from the Allergy Clinic, Phramongkutklao Hospital (Bangkok, Thailand). Healthy subjects were enrolled with no history of allergic disease and medication used. AR subjects with positive skin prick tests (wheal diameter ≥ 6 mm greater than a negative control) to the house dust mite Dermatophagoides pteronyssinus extract (Alk-Abello, Round Rock, TX, USA) were recruited in the study. AR subjects with mite-specific immunotherapy (Alk-Abello) over 2 years were enrolled in the study. A 30-week weekly dose escalation protocol was used with subcutaneous administration of standardized mite mixes, contain glycerin 50% v/v, phenol 0.4% (preservative), sodium chloride, sodium bicarbonate, D. pteronyssinus 5,000 allergy units/mL and Dermatophagoides farinae 5,000 allergy units/mL (Alk-Abello). The AIT subjects were treated with the maintenance dose that had been given at 4-week intervals over 2 years of therapy. Allergy symptoms were recorded using the visual analog scale (VAS) of 100 mm range and the patients marked on a line the point they felt represented their perceptions of their current state. Medication score was scored on a scale of 0-5 (0, not at all; 1, occasionally seldom; 2, occasionally often; 3, almost daily; 4, continuously; 5, continuously with the maximal dose) that included oral anti-histamine, oral decongestant or nasal corticosteroid spray. Subjects who presented with no allergic symptoms (VAS, 0–10 mm) and discontinued medication (medication score, 0–1) were classified as clinical responders to AIT. Subjects who still had allergic symptoms (VAS, 60–100 mm) and could not reduce medication to control allergic symptoms (medication score, 3–5) were classified as clinical non-responders to AIT. The study was performed with the approval by the Royal Thai Army Medical Department Ethics Committee, IRBRTA 1494/2559 and with the patients' written informed consent.
Innate lymphoid cell detection by flow cytometry
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque (Biochrom, Berlin, Germany) density-gradient centrifugation of peripheral venous blood. PBMCs were stained with Fixable Viability Dye eFlour780 (eBiosciences, San Diego, CA, USA) according to the manufacturer's instructions. After the cells were washed with 500 µL of staining buffer (phosphate-buffered saline, 0.5% bovine serum albumin and 2 mmol/L ethylenediaminetetraacetic acid), cells were subsequently stained with antibodies against surface markers. In brief, FITC-lineage marker (CD1a, CD11c, CD34, CD94, CD123, CD303, FcεRIα, CD3, CD4, CD8a, CD14, CD16, and CD19), PE-dazzle 594-c-Kit, PerCP-CY5.5-CD161, PE-Cy7-CD127, CD69-AF700, and BV510-CD45 (all antibodies from Biolegend, San Diego, CA, USA) and AF647-CRTH2 (BD Biosciences, San Jose, CA, USA) were stained at room temperature for 15 minutes. Then, cells were washed 2 times with staining buffer. Cells were analyzed with a Galios (Beckman Coulter, Indianapolis, IN, USA). Flow cytometry data and dot plot graphs were analyzed with the FlowJo v10 software (FlowJo, Ashland, OR, USA).
Allergen-specific re-stimulation
For ILC2s activation, PBMCs were cultured with 1 µg/ml recombinant Der p 1 protein (Prospec, Rehovot, Israel),100 U/mL IL-2 (Sigma-Aldrich, St Louis, MO, USA) and 50 ng/ml IL-33 (Prepro Tech, Rocky Hill, NJ, USA) in complete RPMI medium 1640 supplement with L-Glutamine (cRPMI, ThermoFisher, Waltham, MS, USA), 10% fetal bovine serum (Sigma-Aldrich), and 1% PenStrep (Sigma-Aldrich). Cells were then cultured at 37°C in humidified atmosphere in 5% CO2 for 3 days. After that, cells were stained with Fixable Viability Dye eFlour780 and antibodies as previously described. Mean fluorescence intensity of activation marker (CD69 expression).
Statistical analysis
All data are shown as mean ± standard deviation (SD) of the study groups. One-way analysis of variance and Tukey's multiple comparison tests were used to analyze for comparisons between the groups. Statistical analysis was performed with GraphPad Prism 7.0 Software (GraphPad Software, La Jolla, CA, USA). A P value of less than 0.05 was considered statistically significant.
RESULTS
Subject characteristics
Table details the proportion of male and female enrolled in the study, the mean age, the duration of AR, the frequency of co-morbid allergic diseases, the mean total immunoglobulin E (IgE) levels, the mean VAS and the mean medication score. PBMCs were obtained from 100 adult subjects (28 healthy subjects, 35 AR patients, and 37 AIT patients). Of 37 patients with AIT, 26 were classified as clinical responders and 11 were classified as clinical non-responder to therapy according to VAS and medication score. There are an almost equal sex proportion and the mean age between the groups. We found that the mean total IgE levels were lower in the healthy group compared to AR patients. There was no significant difference in mean VAS and medication score between non-responder AIT and AR patients.
Table
Baseline characteristics of subjects

The frequency of circulating ILCs
The gating strategy of ILCs is shown in Fig. 1. ILCs were gated as CD45+Lin−CD127+CD161+. The expression of CRTH2 and c-Kit were used to identified ILC2 and ILC3 populations. ILC1s were identified as lack expression of CRTH2 and c-Kit. ILC3s NCR+ were characterized by the expression of the natural cytotoxic receptor NKp 44.
Fig. 1
ILCs gating strategy use to describe ILC1s, ILC2s and ILC3s from peripheral blood. Representative flow cytometry dot plot demonstrating ILC gating strategy, ILC were defined as CD45+ Viable Lin−CD127+CD161+. ILCs were characterized into 3 groups based on CRTH2 and c-Kit, ILC1s (CRTH2−c-Kit−), ILC2s (CRTH2+), and ILC3s (CRTH2−c-Kit+). NKp44 in humans is used to gate ILC3s NCR+ (NKp44+) and ILC3s NCR− (NKp44−).
ILC, innate lymphoid cell; NCR, natural cytotoxicity receptor.
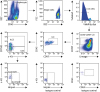
As for frequencies of ILC subsets in patients undergoing HDM immunotherapy, clinical responders showed a higher frequency ILC1s compared to non-responder patients (64.71% ± 10.34% vs.46.53% ± 8.5%, P = 0005; Fig. 1A and B). In this study, we also found that frequency of ILC1s was higher in responder AIT patients than in healthy subjects and AR patients. Moreover, the healthy group showed significantly higher average frequency of ILC1s compared to AR patients (54.7% ± 13.23% vs.37.88% ± 13.65%, P < 0.0001; Fig. 2A and B).
Fig. 2
ILC1s and ILC2s from human peripheral blood. (A) Representative dot plots of ILCs population, and mean of ILC1s and ILC2s. Frequency of ILC1s (B) and ILC2s (C) were expressed as percentages of the ILCs (mean ± SD) in healthy (n = 28), AR (n = 35), non-responder AIT (n = 11), and responder AIT (n = 26). The data were analyzed by analysis of variance and Tukey's multiple comparison test.
ILC, innate lymphoid cell; SD, standard deviation; AR, allergic rhinitis; AIT, allergen-specific immunotherapy.
*P < 0.05, †P < 0.001, ‡P < 0.0001.

ILC2s have recently emerged as play an important role in the development of allergic inflammation. In contrast to the results with ILC1s, subjects with AR which were not treated with AIT revealed significantly higher mean frequency of ILC2s (41.85% ± 13.8%) in their peripheral blood than healthy subjects (22.08% ± 10.5%, P < 0.0001) and responder AIT patients (23.15% ± 8.54%, P < 0.0001; Fig. 2A and C). Notably, subjects who responded to AIT had a lower amount of ILC2s compared to non-responder AIT patients (39.12% ± 4.74%, P = 0.0006) and almost at the same level as healthy subjects (Fig. 2A and C). In addition, the mean frequency of ILC2s was higher in non-responder AIT patients than in healthy subjects, but nearly the same levels compared to AR patients without AIT (Fig. 2 A and C).
To fully characterize the frequency of ILC3s, we measured NCR+ and NCR− population and compared them between the groups. The frequencies of ILC3s NCR+ and NCR− were low in AIT (Fig. 3A). There was a statistically significant difference in the frequency of ILC3s NCR+ between patients who were responder AIT (1.51% ± 0.88%) compared to healthy subjects (3.59% ± 2.45%, P = 0.0009) and AR patients (3.27% ± 2.13%, P = 0.003), but no significant difference compared to non-responder AIT patients (2.31% ± 1.6%; Fig. 3B). However, the average frequency of ILC3 NCR+ in healthy subjects showed no significant difference compared to AR patients (Fig. 3B). Similarly, the results of ILC3s NCR− was significantly lower in the responder AIT patients (9.92% ± 8.43%) compared to healthy subjects (18.26% ± 8.42%, P = 0.002) and AR patients, (18.99% ± 9.05%, P = 0.0005), but there was no significant difference compared to non-responder AIT patients (2.31% ± 1.6%; Fig. 3C). The frequency of ILC3s NCR− was not statistically different between healthy subjects and AR patients. We also analyzed the correlation between clinical severity scores and the frequencies of ILCs. The ILC1s levels showed a negative correlation with VAS (r = −0.68) and medication score (r = −0.65), while ILC2s, ILC3s NCR+ and ILC NCR− showed a positive correlation with VAS (r = 0.62, 0.36, and 0.45, respectively) and medication score (r = 0.56, 0.35 and 0.31, respectively; Supplementary Fig. S1A and B).
Fig. 3
ILC3s NCR+ and NCR− from the human peripheral blood. (A) Representative dot plots and mean of ILC3s NCR+ and NCR−. Frequency of ILC3s NCR+ (B) and NCR− (C) were expressed as percentages of the ILCs (mean ± SD) in healthy (n = 28), AR (AR, n = 35), non-responder AIT (n = 11) and responder AIT (n = 26). (D) Proportions of ILC subsets in each group. The data were analyzed by analysis of variance and Tukey's multiple comparison test.
ILC, innate lymphoid cell; NCR, natural cytotoxicity receptor; SD, standard deviation; AR, allergic rhinitis; AIT, allergen-specific immunotherapy.
*P < 0.01, †P < 0.001.

The ILCs proportion was also investigated (Fig. 3D). The effect of AIT might shift the proportion of ILC2s to ILC1s in HDM AR patients. The responder AIT patients had higher ILC1 to ILC2 proportion compared to healthy subjects. Meanwhile, the non-responder AIT patients had a high ILC2 to ILC1 portion similar to AR patients without AIT. Therefore, our current results show that AIT affected the ILCs proportion in the peripheral blood of HDM AR patients.
AIT suppresses circulating ILC2s activation
The percentage of activated (CD69+) ILC subsets was investigated. At baseline without any stimulation, no significant difference was found in the frequency of activated ILC1s, ILC2s, ILC3s NCR+, and NCR− between the groups (Supplementary Fig. S2A-E). Next, we cultured 65 PBMCs (20 healthy subjects, 20 AR patients, 9 non-responder AIT patients, and 16 responder AIT patients) with recombinant Der p 1 protein, IL-33, and IL-2 to determine the responses of ILC2 activation with allergen specificity. We analyzed the mean fluorescence intensity (MFI) of CD69 expression on ILC2s at baseline compared to that after in vitro stimulation (Fig. 4A). Fold change of activated ILC2s was significantly higher in AR patients compared to healthy subjects (P = 0.0003; Fig. 4B), which confirmed the response of ILC2s in allergic patients with allergen specificity. Interestingly, the response of activated ILC2s was also significantly lower in responder AIT patients compare to AR patients (P = 0.001; Fig. 4B). However, no difference was observed between responder and non-responder AIT patients in terms of fold change CD69 expression. The cytokine levels in the supernatants were measured, but no significant difference between the groups was observed (Supplementary Fig. S3). Therefore, our current results show that AIT suppressed circulating ILC2s activation in responder AIT patients.
Fig. 4
Activated ILC2s decreased in responder AIT patients. (A) Representative overlay histogram graph of CD69 expression on ILC2s between baseline and 3 days after stimulation with the recombinant antigen-specific protein. (B) Fold change in the mean fluorescence intensity of CD69 expression on ILC2s from healthy (n = 20), AR (AR, n = 20), non-responder AIT (n = 9), and responder AIT (n = 16) in response to allergen-specific stimulation. The data were analyzed by analysis of variance and Tukey's multiple comparison test.
ILC, innate lymphoid cell; AIT, allergen-specific immunotherapy; AR, allergic rhinitis.
*P < 0.01, †P < 0.001.

DISCUSSION
The data reported here demonstrates that the frequencies of ILC1s and ILC2s are different between responder and non-responder AIT patients and also different between healthy subjects and AR patients without AIT. Moreover, we also show ILC3s NCR+ and NCR− are lower in responder AIT patients compared to healthy subjects and AR patients. Currently, ILC2s have been shown to play an important role in allergic diseases such as asthma, atopic dermatitis and chronic rhinosinusitis.31 The role of ILC2s in AR is still poorly understood. Increased epithelial pro-allergic cytokines (IL-25, IL-33, and TSLP) that affect activated ILC2s were detected in nasal lavage in HDM AR patients.3233 ILC2s alone without local Th2 cells cannot mediate the symptoms of AR in a mouse model.34 ILC2s represent an abundant alternative source of Th2 cytokines and likely serve to increase and maintain local Th2-driven allergic inflammation.35 In HDM allergy study, circulating ILC2s levels were increased in AR patients and correlated with symptom severity.23 In the same study, they also found higher IL-5 and IL-13 levels that were produced by ILC2s in AR patients than in healthy control.23 To our knowledge, there have been no reports of ILC1s and ILC3s levels in HDM AR patients. In grass pollen allergy, the frequency of circulating IFN-γ+ILC1s was lower in AR patients than in non-allergic subjects, whereas the frequency of circulating ILC3s was not significantly different between AR and non-allergic subjects.22
AIT is effective in selected patients with AR, particularly in those with seasonal rhinitis and perennial allergy caused by house dust mites.353637 The T cell mechanism for AIT induces immune tolerance by immune deviation to a competitive Th1 response through the induction of a suppressive regular T (Treg) cells response and through the induction of anergy or deletion of Th2 cells.3839 Antigen-presenting cells (APCs) present antigens to IL-12-induced, IFN-γ-producing Th1 cells and inhibit the differentiation of Th2 cells.40 An increase in Treg cells leads to a dose-dependent decrease in cytokine production by ILC2s.41 Moreover, an in vitro study showed that recombinant TGF-β or IL-10 suppressed both human and murine ILC2s.42 In addition, human inducible Treg cells suppress syngeneic human ILC2s through the ICOSL to control airway inflammation in a humanized ILC2 mouse model.42
ILC2 is functionally plastic.43 Especially, the IL-12/IL12R-signaling axis is shown to skew human IL-13-expressing ILC2s to IFN-γ and T-bet expressing ILC2s.44 Moreover, IL-4 could reverse IL-12-induced skewing of ILC1s to ILC2s.1444 This might be a possible mechanism that explains who is a responder AIT with shifted circulating IL-4-producing Th2 cells to IFN-γ-producing Th1 cells related to shifted circulating ILC2s to ILC1s. Our results show similarities with the previous HDM immunotherapy in AR patients study, the levels of ILC2s in the peripheral blood of immunotherapy group were significantly reduced compared to the untreated group, but were not significantly different from healthy controls.30 AIT significantly inhibits Th17-mediated inflammation in HDM AR patients.45 However, no apparent evidence of the correlation between ILC3s and AIT was reported. In the mouse model, IL-10-secreting regulatory ILCs suppress the activation of ILC3s.46 As for Th-cell commitment, cytokine-mediated conditioning as well as epigenetic and transcriptional regulation may account for changes in ILC subset-determining transcription factors and cytokines.47 This might also explain our results that show a decrease in the frequency of ILC3s in responder AIT patients.
In our study, responder AIT patients showed a lower response of activated ILC2 after stimulated with allergen-specific than AR patients. We used recombinant Der p 1, IL-33, and IL-2 to measure activated ILC2s responses with allergen specificity. ILCs do not express antigen receptors, but they can be activated by several factors.31 We found that only recombinant Der p 1 did not show strong stimulation to ILC2s in an in vitro study (data not shown). IL-33 is used to activate ILC2s.48 IL-33 is present in epithelial cells, endothelial cells and macrophages that bind to ST2 receptor on ILC2s and induce IL-13 production.43 IL-2 is essential for immature and mature T cell survival.49 IL-2 can also activate ILC2 and enhance IL-10-producing ILC2s.3150 Besides cytokines, lipid mediators are also able to stimulate ILC2 function.46 In our pilot study, we used 1 µM PGD2 to activate ILC2s, but we did not find any difference in CD69 expression after stimulation between healthy subjects and allergic patients (data not shown). As shown in this study, a combination of IL-33 and allergen-specific protein can stimulate activated ILC2s and then show a discriminatory effect between healthy subjects and allergic patients. However, we failed to demonstrate that this method can be used to differentiate between responders and non-responders to AIT therapy.
Because our study was a cross-sectional study, our results were not longitudinal to monitor the frequency of ILCs compared from baseline to a few years after AIT. We could not conclude that AIT could modulate circulating ILC blood frequencies during the course of the therapy. Additionally, we did not measure either the cytokine producing-ILCs, transcription factors used to identify ILC subsets, or levels of Th2 cytokines in serum. In our next study, we will attempt to demonstrate the dynamic change in the frequencies of ILC subsets during AIT.
In conclusion, our study presented evidence that increased circulating ILC2s are involved in AR pathological mechanism. The ILC2 response was reduced in AR patients undergoing AIT. AIT might be affected by the ILC populations by skewed ILC2s to ILC1s and decreased ILC3s. The responder AIT patients and healthy controls show a similar ILC2: ILC1 proportion. The clinical relevance of ILC2s and AR suggest that ILC2s can be a potential biomarker for the therapeutic effect of AIT in AR patients.
 XML Download
XML Download