

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Osseointegrated implants used to treat partially or fully edentulous patients have shown predictable long-term success and survival rates. However, reports describing post-implantation complications and implant failures have recently become more frequent. Implant failures can be categorized as early and late failures. Early failures are often caused by problems in the surgical operation itself or by implant- and patient-related factors before functional loading is applied. Late failures occur after prosthetic loading, and are usually caused by overloading or peri-implant disease. Bacterial infections are the major cause of peri-implant disease, which is classified as peri-implant mucositis and peri-implantitis [1]. Among the more than 500 species of microorganisms that exist in the oral cavity, certain anaerobic, gram-negative species often aggravate periodontal disease. Paster et al. [2] established that Porphyromonas gingivalis, Prevotella intermedia, Aggregatibacter actinomycetemcomitans, Tannerella forsythia, and Treponema denticola play a critical role in deep periodontal pockets. These species can affect not only the periodontal pockets involved in periodontitis, but also peri-implant tissue. Bacteria colonize the peri-implant sulcus within two weeks of implant placement. Further plaque accumulation in the site leads to peri-implant infection and the advancement of peri-implant disease [1]. The bacterial species found in healthy peri-implant sulci are very similar to the subgingival bacteria in healthy periodontal tissue, and the bacterial species found in cases of peri-implant disease are very similar to the subgingival bacterial complexes found in chronic or recurrent periodontitis patients [3]. P. gingivalis and A. actinomycetmcomitans have been found in the buccal gingival sulci of implant abutments [4]. As transmission of microbes is common within the oral cavity, the condition of the residual teeth influences the microbial composition of early plaque around implants [5]. Mombelli et al. [5] reported that more pathogenic bacteria were detected in residual teeth with a history of periodontal disease than in teeth without such a history.
One of the key periodontal pathogens, P. gingivalis is gram-negative, black-pigmented, and anaerobic [6]. P. gingivalis is not only responsible for periodontal disease in the natural dentition, but it also associated with peri-implant tissue destruction. Salcetti et al. [7] reported a higher detection rate of P. gingivalis, T. forsythia, and T. denticola species in failed implants in comparison with healthy implants. Rutar et al. [8] reported a significant relationship between peri-implant probing depth and the detection of P. gingivalis. Botero et al. [9] compared the bacterial species found in healthy peri-implant tissue with those found in tissue with peri-implant disease, and P. gingivalis was only found in tissue with peri-implant disease.
The virulence factors of P. gingivalis include fimbriae, its capsule, collagenase, and gingipains [10,11]. In particular, the fimbriae of P. gingivalis play a critical role in adherence to the host cell, facilitating bacterial invasion and infection [10,12,13]. They also promote early plaque formation and regulate plaque maturation [14]. Various inflammatory cytokines (IL-1α, IL-β, IL-6, TNF-α) expressed by the fimbriae facilitate alveolar bone resorption [15]. Lee et al. [16] were the first to report mutations of the FimA protein, and Nakagawa et al. [17] identified six different genotypes (types I–V, Ib) of the fimA gene, which encodes fimbrillin, a subunit of the fimbriae, according to nucleotide sequences. Nakagawa et al. [13] reported that type II fimA was capable of more efficient attachment to host cells and cell invasion, thus playing a more important role in the pathogenicity of periodontal disease than other genotypes. However, Umeda et al. [18] found no significant differences in epithelial attachment and invasion among the different genotypes. Amano et al. [19] reported that P. gingivalis with type I fimA was closely associated with periodontally healthy individuals. Nagano et al. [20] found the expression of fimA to be positively related to plaque accumulation for all genotypes, with type I fimA showing an especially close relationship to plaque formation. The prevalence of P. gingivalis fimA genotypes in peri-implant sulci was studied by Shin et al. [21] and Seo et al. [22], and P. gingivalis type II fimA was found to be significantly correlated with peri-implantitis.
However, as type Ib fimA shares 97.1% and 77.5% of its nucleotide sequence with type I and type II fimA, respectively, cross-hybridization is likely to occur during polymerase chain reaction (PCR) analysis [17,23]. Type I and Ib fimA genotypes can be discriminated with RsaI-digestion [17], but the possibility of false positive detection with PCR using conventional primers has been reported [17,23].. In order to overcome this problem and increase the accuracy of sequencing, Moon et al. [24] suggested a new primer that is specific for the DNA of type II fimA.
The purpose of this study was to evaluate the relationship between the condition of peri-implant tissue and the distribution of P. gingivalis fimA genotypes, using new primers in Korean subjects.
MATERIALS AND METHODS
Study population
The distribution of P. gingivalis fimA genotypes in peri-implant sulci was studied in a group of patients with a history of implant placement who visited the Department of Periodontology, Kyung Hee University Dental Hospital, Republic of Korea from January 2007 to November 2011. The exclusion criteria included patients with a history of systemic or local use of antimicrobials three months prior to the study and patients with known systemic conditions that could influence their periodontal status. The control group consisted of sound implants with a peri-implant probing depth of 5 mm or less with no bleeding on probing. Test group I consisted of implants with a peri-implant probing depth of 5 mm or less with bleeding on probing, while test group II consisted of implants with a peri-implant probing depth of more than 5 mm with bleeding on probing. This study was approved by the Ethical Committee of the School of Dentistry, Kyung Hee University (KHUSD IRB2009-02).
Plaque sampling in peri-implant sulci and DNA isolation
Supragingival plaque was removed prior to subgingival plaque sampling. Sampling sites were protected from saliva influx with sterile cotton pellets and compressed air. Sterile paper points were inserted into two deepest pockets of the implants for 30 seconds and removed. The paper points were collected in a sterile tube with 1 mL of sterile phosphate-buffered saline (pH 7.4). Bacterial genomic DNA was extracted from the plaque samples using the InstaGeneTM Matrix kit (Bio-Rad Laboratories, Hercules, CA, USA) according to the manufacturer's instructions. The InstaGene matrix suspension was added to the plaque samples, and the mixture was incubated at 56°C for 15 minutes and boiled at 100°C for eight minutes. After centrifugation, the DNA-containing supernatant obtained was subjected to PCR amplification.
Polymerase chain reaction
The presence of P. gingivalis in peri-implant sulci was confirmed using DNA extracted from the plaque samples by PCR. Certain DNA sequences in acquired plaque samples were identified using universal primers, based on P. gingivalis-specific 16S rRNA sequences, and analyzed further to differentiate their fimA genotypes using the six sets of fimA type-specific primers. The primer newly suggested for type II fimA by Moon et al. [24] was used (Table 1). PCR amplification was performed in a total volume of 20 µL containing 10 µL of PCR Pre-Mix (STD02-M50h; SolGent, Korea), with 0.5 µM of each primer and 5 µL of the template DNA solution in sterile distilled water. DNA amplification was performed with a thermal cycler (Model 9700; Applied Biosystems, Branchburg, NJ, USA): an initial denaturation was performed at 95°C for five minutes; followed by 30 cycles at 94°C for 30 seconds, 55°C for 30 seconds, and 72°C for 30 seconds; and a final extension at 72°C for seven minutes.
Table 1
Oligonucleotide primers used in this study

| Primer set | Primer sequence (5'-3') | Size (bp) | Reference |
|---|---|---|---|
| Universal | F: AGAGTTTGATCMTGGCTCAG | 315 | Tamura et al. (2005)[39] |
| R: CTGCTGCSYCCCGTAG | |||
|
P. gingivalis 16S rRNA |
F: TGTAGATGACTGATGGTGAAAACC | 197 | Amano (1999)[7] |
| R: ACGTCATCCCCACCTTCCTC | |||
| Type I | F: CTGTGTGTTTATGGCAAACTTC | 392 | Amano (1999)[7] |
| R: AACCCCGCTCCCTGTATTCCGA | |||
|
Type II (new) |
F: GCATGATGGTACTCCTTTGA | 292 | Moon et al. (2011)[25] |
| R: CTGACCAACGAGAACCCACT | |||
| Type III | F: ATTACACCTACACAGGTGAGGC | 247 | Amano (1999)[7] |
| R: AACCCCGCTCCCTGTATTCCGA | |||
| Type IV | F: CTATTCAGGTGCTATTACCCAA | 251 | Amano (1999)[7] |
| R: AACCCCGCTCCCTGTATTCCGA | |||
| Type V | F: AACAACAGTCTCCTTGACAGTG | 462 | Nakagawa et al. (2000)[40] |
| R: TATTGGGGGTCGAACGTTACTGTC | |||
| Type Ib | F: CAGCAGAGCCAA AAACAATCG | 271 | Nakagawa et al. (2002)[18] |
| R: TGTCAGATAATTAGCGTCTGC |
![]()
P. gingivalis-positive specimens were classified according to six different types of specific primer sets. The PCR products were electrophoresed on 1.8% agarose gel (Figure 1). Type I and Ib fimA genotypes were further categorized by RsaI-digestion (Figure 2).
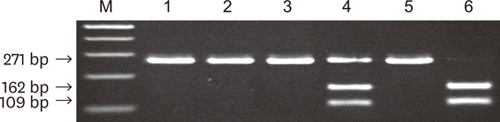
 | Figure 1Electrophoresis of amplification products on a 1.8% agarose gel. Lines 1–6 are type I, type Ib, type II (new), type III, type IV, and type V. M, molecular weight marker.
|
 | Figure 2Detection of type Ib fimA by polymerase chain reaction amplification and RsaI digestion. Lanes 1–3 comprise fimA amplicons from a mixed culture of P. gingivalis strains (ATCC33277 for type I fimA and HG1691for type Ib fimA), the pure culture of strain ATCC33277, and the pure culture of strain HG1691, respectively, using type Ib primers. Lanes 4–6 show the amplicons of lanes 1-3 digested with RsaI. M, molecular weight marker.
|
Statistical analysis
The power of the goodness of fit or the chi-square independence test is given by 1−β−Fdf, λ
(xcrit
), where F is the cumulative distribution function for the noncentral chi-square distribution χ
2(df); xcrit
is the χ
2(df) critical value for a given value of alpha; and λ□= w
2
n is the noncentrality parameter, where w is the φ□effect size and n is the sample size. Based on a formula reflecting a medium effect size (w = 0.3) [25], we calculated that the sample size required per group was 36 and that the total sample size required was 108, at an alpha of 0.05 and a power of 0.8. Assuming a prevalence of P. gingivalis ranging between 55% and 95% in Korean adults [26], we estimated that a total of 114–198 subjects would be required.
The comparative frequencies of P. gingivalis in the control and test groups and the frequency of each fimA genotype were analyzed using the chi-square test or Fisher’s exact test. In all statistical analyses, P-values < 0.05 were considered statistically significant. All statistical calculations were performed using SPSS 18.0 (SPSS Inc., Chicago, IL, USA).
RESULTS
The study consisted of 184 subjects: 88 men (age range, 32–76 years; mean age, 57.0±11.4 years) and 96 women (age range, 29–72 years; mean age, 54.2±10.4 years). Among 248 plaque samples, P. gingivalis was detected in 236 samples (177 subjects; mean age, 55.0±9.5 years), with an infection rate of 95.2%. Ultimately, 85/91 (93.4%) samples from the healthy control group, 96/101 samples (95.0%) from test group I, and 55/56 samples (98.2%) from test group II were included in the study. Of the 177 P. gingivalis-positive subjects, 61 were in the control group, 70 were in test group I, and 46 were in test group II. A single plaque sample was collected from 133 subjects, while two or more samples were collected from 44 subjects. The distribution of sample collection sites is presented in Table 2. More than 70% of the samples were from implants in posterior sites.
Table 2
Distribution of sample collection sites

![]()
Table 3 shows the distribution of P. gingivalis according to the fimA genotype detected in plaque specimens. In the control group, type I was detected in 11 specimens (12.9%), type II in 51 specimens (60.0%), type III in two specimens (2.4%), type IV in nine specimens (10.6%), and type Ib in five specimens (5.9%). No fimA genotype was detected in 23 specimens (27.1%). In test group I, fimA type I was found in 10 specimens (10.4%), type II in 62 specimens (64.6%), type III in seven specimens (7.3%), type IV in 14 specimens (14.6%), and type Ib in nine specimens (9.4%). Nineteen specimens (19.8%) showed no fimA genotype. Test group II showed fimA type I in nine specimens (16.4%), type II in 32 specimens (58.2%), type III in 0 specimens (0.0%), type IV in nine specimens (16.4%), and type Ib in 12 specimens (21.8%). No genotype was detected in nine specimens (16.4%). Type V was not detected in any group. It was not the case that each specimen expressed a single fimA genotype; in some cases, two or more genotypes were detected together. Two genotypes were detected in seven specimens (8.3%) in the control group, 13 specimens (13.5%) in test group I, and eight specimens (14.5%) in test group II. More than three genotypes were detected in four specimens (4.7%) in the control group, six specimens (6.3%) in test group I, and three specimens (5.5%) in test group II. The amount of specimens with a single genotype in all three groups was similar, and the differences among groups in this regard were not statistically significant. The most frequently detected P. gingivalis fimA genotype was type II in all groups. None of the fimA genotypes in test group I exhibited a distribution significantly different than was observed in the control group. Test group II showed a statistically significant difference regarding type Ib fimA.
Table 3
Distribution of the six fimA genotypes in P. gingivalis - positive samples

a)Statistically significant differences between the control group and test group II (P<0.05).
b)The chi-square test
c) Fisher’s exact test
![]()
The distribution of fimA genotypes according to pocket probing depth of the patients in the test groups showed a 7.3% detection rate of the type III genotype in patients with a probing depth of 5 mm or less, which was significantly higher than the detection rate in patients with a probing depth greater than 5 mm. Type Ib was detected in 21.8% of specimens from patients with a probing depth greater than 5 mm, which was significantly higher than the detection rate in specimens with a probing depth of 5 mm or less (Table 3).
DISCUSSION
This study examined the prevalence of P. gingivalis fimA genotypes in plaque samples from the peri-implant sulci of Korean patients by PCR, using a new primer for type II fimA. The control group showed a P. gingivalis infection rate of 93.4%, with rates of 95.0% and 98.2% for test groups I and II, respectively. These differences were not statistically significant. The presence of P. gingivalis in general may not be related to peri-implant disease; instead, it is possible that bacterial colonies with a specific genotype influence peri-implant tissue destruction. Therefore, not all bacterial colonies of the same species necessarily show the same pathogenicity. Instead, the severity and progression of the disease may be dependent on the specific genotype of the bacterial colonies.
Previous studies of the distribution of P. gingivalis fimA genotypes were mostly based on patients with periodontitis. P. gingivalis type II fimA was the most frequently detected, followed by type IV, as in studies of Japanese and Chinese patients with chronic periodontitis [6,17,19,27]. Type II and type Ib were detected most frequently in Brazilian [28] and Spanish [29] populations. Although differences among studies exist, type II fimA has been found to be the most closely related to chronic periodontitis, corresponding to the results of the present study based on a Korean population sample, in which the highest detection rate of type II fimA was observed in patients with peri-implant lesions. However, unlike the findings of previous studies that reported a low detection rate of type II fimA in healthy teeth [17,27], we also frequently found type II fimA around healthy implants. This can be interpreted in two possible ways: either healthy implants with type II fimA have the potential to develop peri-implantitis in the future or type II fimA is not a risk indicator for implants, unlike for natural teeth.
In this study, type Ib fimA was found to be detected significantly more frequently in test group II than in the other groups. P. gingivalis with type Ib fimA plays a critical role in peri-implant tissue destruction and is a potential risk indicator for peri-implantitis.
The fimA genotypes in implants with inflammatory lesions were analyzed according to probing depth. Type Ib was detected in 8.9% of samples with a pocket depth of 5 mm or less, versus in 21.4% of samples with a pocket depth greater than 5 mm, which was a statistically significant difference. Therefore, the presence of type Ib fimA was related to deepening of the probing depth over the course of the progression of implant disease. Type III was only present in patients with a probing depth of 5 mm or less, which corresponds to previous reports in which type III was observed in periodontally healthy individuals and/or in sites with a low probing depth [19,23,27]. Nakano et al. [30] subcutaneously injected variable P. gingivalis fimA genotypes and compared the inflammatory response, finding that type III showed the weakest inflammatory response.
Unlike previous reports, the present study found noteworthy results regarding type Ib fimA relative to type II. Several potential reasons exist for this discrepancy. First, unlike preceding studies, this study used new primers to prevent the false negative or false positive detection of type II fimA. The fimA genotype was originally classified into five types (types I to V), before Nakagawa et al. [17] defined type Ib. Several studies may have mistakenly detected type Ib as type II due to accidental cross-hybridization, meaning that the relative risk of type II could have been overestimated [26]. In order to reduce the likelihood of such errors, Moon et al. [24] developed a new primer that was shown to be effective in promoting accurate detection of type II fimA. Recent studies on the prevalence of each fimA genotype of P. gingivalis in the Korean population using new primers for type II have shown a high prevalence of types II, Ib, and IV fimA in periodontitis patients. In particular, the correlation between chronic periodontitis and type Ib fimA was found to be the most significant [26]. This corresponds to the finding of the present study that type Ib fimA was the genotype most significantly related with peri-implant inflammatory lesions. Second, the results of a study evaluating the fimA genotypes of P. gingivalis in peri-implant sulci may be different from those reported in studies of natural teeth. Subramani et al. [31] demonstrated variation in the composition of bacterial plaque around implants and natural teeth caused by the fact that they have different surface roughness and surface energy. According to Koyanagi et al. [32], a more complicated microbial complex of plaque existed in peri-implantitis sites than in periodontitis sites, and periodontal pathogens were detected less frequently. Environmental differences between peri-implant and periodontal sulci account for the variability of the P. gingivalis fimA genotypes associated with periodontitis and peri-implantitis. Third, fimbriae are not the only virulence factors of P. gingivalis, and the presence of other virulence factors may have affected the findings of previous studies. Representative virulence factors include encapsulation (K1–K6 serotypes) and gingipains. Gingipains are a type of proteolytic enzyme, and can be further classified into arginine-specific types A, B, and C and lysine-specific types I and II. Laine and van Winkelhoff [33] reported that encapsulated P. gingivalis was associated with the formation of more severe periodontal abscesses. Imamura [11] concluded that gingipain functions as a virulence factor by inhibiting the host immune response, increasing the tendency for bleeding, and promoting inflammation, thus aggravating periodontal disease. Therefore, periodontal disease may develop through a complex and combined response to various virulence factors not associated with P. gingivalis fimA genotypes. Fourth, variability may exist across racially and geographically diverse populations. Rylev and Kilian [34] found a diverse distribution of the genotypes of the periodontal pathogens P. gingivalis and A. actinomycetemcomitans in different races, which in turn influenced the prevalence of periodontitis. Many studies drawing on subjects from distinct ethnic groups have shown different fimA genotypes to be associated with virulence. The study of van der Ploeg et al. [35] was based on periodontitis patients in Norway and Switzerland, and type IV was detected more frequently than type II. Additionally, Beikler et al. [36] reported no correlation between the severity of periodontal disease and P. gingivalis fimA genotype in a study based on Caucasians. The higher prevalence of type I fimA reported in this study in comparison to a previous study including Asian subjects may be explained as a result of differences in the geographic location. Thus, the novel results of this study regarding the distribution of the type Ib genotype may only be pertinent for the Korean population.
In the present study, a single P. gingivalis fimA genotype was detected in approximately 60% of the control and test groups, and two or more types were also found in a subset of the samples. The multiple genotype occurrence rate in the test groups was approximately 20%, which is similar to the results of several previous studies (23%-26%) [17,19,23]. The prevalence of P. gingivalis with unclassified genotypes was high compared to the findings of Amano et al. [6] and Nakagawa et al. [17]. Hayashi et al. [37] explained this variability as being due to sampling methods. The sterile paper points used to acquire plaque samples in peri-implant sulci may not be as effective as the direct collection of plaque using curettes, meaning that fewer specimens of P. gingivalis were obtained than would have been possible using curettes. This may have contributed to the fact that we found a higher number of P. gingivalis genotypes that we were unable to classify. Additionally, Perez-Chaparro et al. [38] pointed out the limitations of PCR in discriminating variance within the fimA genotype of P. gingivalis. Little is understood about the influence of unclassified P. gingivalis groups on peri-implantitis. The presence of unclassified genotypes suggests the possibility that some fimA genotypes remain to be discovered and classified, and future studies are expected on this topic.
This study found that a higher prevalence of P. gingivalis with type Ib fimA was significantly associated with peri-implant disease progression. Further studies of the fimA genotype distribution of P. gingivalis in a wide range of ethnic groups are necessary, and would be facilitated by increasing the sensitivity of PCR and lowering the rate of unclassified P. gingivalis genotypes through the use of new primers.
 XML Download
XML Download