

 PDF
PDF ePub
ePub Citation
Citation Print
Print


More than 250 million people worldwide carry hepatitis B virus (HBV). These people are prone to developing liver failure and diseases such as cirrhosis and hepatocellular carcinoma. Although vaccination and anti-HBV therapies are available, HBV infection remains a major global health concern because clearing the virus from chronic HBV carriers is difficult using current treatment methods.
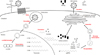
HBV is a DNA virus with reverse transcriptional activity, belonging to the same family (Hepadnaviridae) as woodchuck hepatitis virus and duck hepatitis B virus (Figure 1). The infectious HBV virion, also known as the Dane particle, has a globular structure (approximately 42 nm in diameter) that contains the nucleocapsid [12]. The HBV capsid comprises 120 copies of core protein dimers [34] enclosed within an envelope, which consists of a lipid bilayer membrane derived from the host cell. Embedded in this bilayer are three viral surface proteins (small surface protein [SHB], middle surface protein [MHB], and large surface protein [LHB]). The nucleocapsid harbors a partially double-stranded HBV genome (approximately 3.2 kB) known as relaxed-circular DNA (rcDNA) and the viral polymerase (P protein), which is cross-linked covalently to the rcDNA [56].
A major cause of chronic HBV infection is covalently closed circular DNA (cccDNA), which forms a minichromosome in the host cell nucleus and serves as a template for viral transcription by host RNA polymerase [7]. Therefore, strategies targeting cccDNA may form the basis for developing novel antiviral agents. Recent studies show that interferon-α or APOBEC3 proteins reduce the amount of cccDNA [89]; however, this is not sufficient for complete elimination of cccDNA from host hepatocytes.
HBV infection and development of hepatitis are complex responses involving many factors. Thus, limited investigations have been undertaken using in vitro systems. The host range of HBV is narrow; therefore, few primates are susceptible to HBV infection. Among these, only chimpanzees are immunocompetent and fully susceptible to human HBV [1011]. They develop both acute and chronic HBV infection, along with hepatitis and immune response that are similar to those in humans [12]. However, ethical constraints, large size, and high cost make chimpanzees unsuitable animal models. Tupaias can be infected with HBV, but infection is mild and transient, and viral replication is limited [13]. In addition, these animals are large and hard to handle. Thus, small, easy to handle, and well-defined animal models such as mice are required. Several mouse models of HBV have been established, and novel mouse models that enable global assessment of the viral infection process and efficacy of antiviral agents are being developed continuously.
The HBV Life Cycle
Entry
The HBV virion attaches to hepatocytes via weak interaction between the preS1 domain of the LHB protein on the HBV envelope and heparin sulfate proteoglycans (HSPGs) on the surface of hepatocytes [14]. The sodium taurocholate co-transporting polypeptide (NTCP) on the host cell surface, which is a receptor for HBV entry [15], allows the HBV virion to pass through the plasma membrane via clathrin-mediated endocytosis [16]. However, the detailed molecular mechanisms underlying the entry process are still not fully understood.
Uncoating
After endocytosis, the nucleocapsid is released from the viral envelope and transported to the nucleus. The underlying process is unclear; however, one possible mechanism is that the arginine-rich C-terminal domain of the core protein may provide a transport signal to a nuclear pore. Also, the nucleocapsid is degraded at the nuclear pore complex, whereupon it releases the rcDNA, which then enters the nucleus [17].
Formation of cccDNA
In the nucleus, rcDNA is transcribed to cccDNA. First, HBV polymerase is released from the 5' terminus of the minus rcDNA strand and the RNA primer is removed from the 5' terminus of the plus strand. Host DNA polymerase κ (POLK) and other DNA polymerases fill the gap in the plus strand [18]. The host DNA repair machinery, such as tyrosyl-DNA phosphodiesterase 2 (TDP2), plays a role in this process [19], although other as-yet-unknown host cellular factors are also involved. The cccDNA is maintained in the nucleus of host hepatocytes as a stable episome, which forms a minichromosome by associating with histones and nonhistone proteins [7].
Transcription and translation
The cccDNA serves as a template for transcription of viral mRNAs by host cellular RNA polymerase 2 [2021]. There are four different HBV mRNAs, which differ in length. The longest mRNA, a 3.5 kB pregenomic RNA (pgRNA), encodes the polymerase protein and the precore/core protein. A 2.4 kB preS mRNA encodes the LHB and MHB proteins, and a 2.1 kB S mRNA encodes the SHB protein. In addition, a 0.7 kB X mRNA encodes the X protein. Transcription of these four transcripts initiates at distinct promoters, but all have overlapping 3' termini. Four promoters and two enhancers play roles in transcription: the core, PreS, S, and X promoters, and enhancers 1 and 2. All four transcripts have 5' cap structures and 3' poly-A tails.
Packaging and synthesis of viral genomic DNA
pgRNA serves as a precursor for synthesis of HBV genomic DNA. Encapsidation is triggered by binding of HBV polymerase to the ε-stem loop structure of the pgRNA 5' terminal region in the cytoplasm [222324]. Because pgRNA is terminally redundant, the stem loop structure is also present at the 3' terminus; however, the polymerase also recognizes the 5' cap structure [25]. Therefore, only the 5' ε-stem loop can serve as an encapsidation signal. After binding the polymerase protein, core proteins are recruited for capsid assembly, and pgRNA and polymerase are encapsidated. Next, using pgRNA as a template, the minus strand of rcDNA is synthesized by the reverse transcriptase activity of HBV polymerase [26]. The N-terminal TP domain of HBV polymerase has a tyrosine (Y63) residue that acts as a protein primer for polymerization of minus-strand DNA [627]. After this step, most of the pgRNA template is degraded by the RNase H domain of HBV polymerase, leaving only a small fragment of RNA. The remaining RNA fragment is used as a primer for synthesis of plusstrand DNA [28], resulting in rcDNA synthesis in the nucleocapsid.
Assembly and secretion
The nucleocapsid moves to the endoplasmic reticulum (ER), where the surface proteins (SHB, MHB, and LHB) are embedded into the membrane [2930]. Enveloped HBV (the Dane particle) is formed in the lumen of the ER and released from the cell through secretory pathways and budding. In addition, two kinds of subviral particle, spherical and filamentous, are secreted from the cell. These do not have a viral nucleocapsid and comprise the viral envelope itself.
Murine Models of HBV
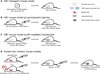
The HBV transgenic mouse
Initially, transgenic mice expressing each viral protein were constructed [3132333435]. These mice were used to evaluate the virology and oncogenic potential of each viral protein. Thereafter, transgenic mice expressing the complete HBV replicon were generated; this model produces infectious virions [36] and made it possible to test the efficacy of antiviral agents such as HBV inhibitors, small interfering RNAs (siRNAs), and cytokines in vivo [37383940]. Since HBV transgenic mice are immune tolerant and HBV pathogenesis proceeds via host adaptive immune responses, the mice do not suffer hepatitis/liver injury or clear the virus. This means that HBV viral proteins or HBV itself is not cytopathic; in addition, HBV-specific cytotoxic T lymphocytes (CTLs) cause acute liver disease when adoptively transferred to these HBV transgenic mice [41]. Thus, studies examining the mechanism(s) underlying infection or cccDNA synthesis are not suited to this model. Since the murine hepatocytes do not express HBV-specific receptors, making intrahepatic infection or spread of HBV impossible, no cccDNA is formed within murine hepatocytes. The inability to clear HBV means that alternative immunocompetent mouse models are required.
Mouse model developed by hydrodynamic injection of HBV replicons
This mouse model is based on immunocompetent mice; therefore, pathogenesis and host immune responses against HBV can be observed [4243]. The HBV replicons are transferred to the mouse liver by injecting a large volume of naked DNA into the tail vein. This DNA is transiently expressed by hepatocytes. Viral replication remains at high levels until Day 7 post-transfection, at which time it starts to decline as HBV-specific antiviral antibodies and CTLs begin to emerge; these are key effectors for viral clearance, which is supported by CD4+ T cells and natural killer cells [42]. This mouse model is used to test antiviral agents such as short hairpin RNAs (shRNAs) and HBV genomespecific guide RNA (gRNA)-mediated clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9 systems [4445]. Recently, the model was used for research into recombinant cccDNA (rcccDNA), which is derived from hydrodynamic-injected HBV minicircle DNA and mimics cccDNA [46].
Mouse model developed by adeno-associated virusmediated transduction of HBV replicons
This model, which is based on viral transduction by intravenously injected adeno-associated virus (AAV), allows more efficient and homogeneous transduction of the liver than the hydrodynamic injection model [4247], in which only 5–10% of hepatocytes are transfected; indeed, more than half of murine hepatocytes in the AAV model express HBcAg [47]. In contrast to the hydrodynamic injection model, this model mimics chronic HBV infection [47]. AAV infection does not induce some types of immune response, and the murine immune system is tolerant to HBV antigens; therefore, HBV persistence is established in injected immunocompetent mice. Due to immune tolerance to HBV, this mouse model can be used to develop anti-HBV vaccines [4748].
Human liver chimeric mouse models
Since mice are not a natural host for HBV, it is not possible to study the entire infection process (from viral entry to cccDNA synthesis and intrahepatic spread) in transgenic or HBV replicon-delivered mice. Therefore, a human liver chimeric mouse model is required.
The first such model was the trimera mouse model, which was established by transplanting HBV-infected human liver fragments under the kidney capsule of lethally irradiated immunodeficient mice [49]. This mouse model has been used to test several antiviral agents; however, since transplanted xenogeneic human hepatocytes do not remain intact for a long time, other models have been developed in which human liver cells are stably engrafted.
The most widely used human liver chimeric mouse model is the albumin-urokinase-type plasminogen activator (alb-uPA)/severe combined immunodeficiency (SCID) mouse [50]. When driven by a liver-specific albumin promoter, uPA is overexpressed and results in subacute liver failure [51]. After backcrossing alb-uPA mice with immunodeficient mice (e.g., SCID or recombination activating gene 2 [RAG2] knockout mice), human hepatocytes are introduced via intra-splenic injection. Next, the small numbers of hepatocytes that reach the injured liver proliferate and repopulate it. The engrafted human hepatocytes are then maintained stably [52]. However, this mouse model has several drawbacks, including infertility, potential fatal bleeding, and kidney disorders [5354]. An alternative human liver chimeric mouse model is the triple knockout FAH-/-RAG2-/- IL2RG-/- (FRG) mouse [55]. This model is generated by crossing fumarylacetoacetate hydrolase (FAH) knockout mice with double immunodeficient RAG2-/-interleukin 2 receptor γ chain (IL2RG)-/- mice. FAH is an enzyme essential for tyrosine metabolism; knocking out FAH leads to accumulation of fumarylacetoacetate in hepatocytes. This compound is toxic to hepatocytes and causes injury [56]. Accumulation of toxic metabolites can be prevented by administration of 2-(2-nitro-4-trifluoro-methylbenzoyl)-1,3-cyclohexanedione (NTBC) prior to transplantation. NTBC is withdrawn upon intra-splenic injection of human hepatocytes. The injected human hepatocytes then repopulate the mouse liver and persist over a period of 6 months [57]. Because human liver chimeric mice are susceptible to HBV infection and cccDNA is formed in transplanted hepatocytes, these models can be used to study viral infection, spread, and the nature of cccDNA. However, immunodeficiency makes these mice unsuitable for study of immune responses induced by HBV or vaccine development.
Conclusions
Here, we discuss the current knowledge base regarding the HBV life cycle and available mouse models (Figure 2 and Table 1). Each step of HBV infection can be targeted by compounds such as mycludex B (an entry inhibitor), APOBEC3 protein or CRISPR/Cas9 (cccDNA inhibitors), siRNAs (viral mRNA inhibitors), entecavir or tenofovir (reverse transcription inhibitors), and BAY 41-4109 (a core protein assembly inhibitor) [9373840444558596061]. However, complete elimination of cccDNA, a key treatment for chronic hepatitis B patients, has not yet been achieved. Development of transgenic and hydrodynamic injection or AAV transduction-mediated replicon delivery model mice allows elucidation of the mechanism(s) underlying the viral life cycle. Also, human liver chimeric mouse models allow examination of viral infection pathways, cccDNA formation, and their sequelae. However, these human liver chimeric mice are immunodeficient. Therefore, to overcome this limitation, immunocompetent human liver chimeric mouse models equipped with human livers and human immune systems have been developed. Examples include albumin promoter-driven-FK506-binding protein-caspase 8 fusion protein (AFC8)-CD34+ human hematopoietic stem cell (hu HSC)/hepatocyte progenitor (Hep) mice and HLA-A2 (A2)/non-obese diabetic SCID IL2RG-/- (NSG)-hu HSC/Hep mice [6263]. However, these mice do not harbor a complete human immune system. Studies of immunopathogenesis, development of novel vaccines, and more comprehensive analyses are feasible if robust dual humanized chimeric mouse models can be generated. This will be one of the most important contributions to HBV research.

 XML Download
XML Download