

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The incidence of airway inflammation has been rising gradually in recent years owing to increases in the atmospheric levels of various toxic materials including pollutants, chemicals, cigarette smoke, and allergens. In particular, chronic obstructive pulmonary disease (COPD), a respiratory disorder characterized by airway inflammation, has become the third leading cause of mortality in the USA [1]. Cigarette smoke (CS) is considered to be a primary risk factor for COPD, leading to airflow limitation and emphysema by inducing airway inflammation [23]. Because CS contains chemicals such as oxidants, aldehydes, nicotine, and carbon monoxide, the lung tissues of smokers have elevated levels of inflammatory mediators, such as chemokines and proinflammatory cytokines, which are released by the lung epithelial cells [45]. These mediators eventually accelerate airway inflammation via increased recruitment of inflammatory cells, particularly neutrophils and macrophages [6].
Neutrophils contribute to the loss of lung function and destruction of normal alveolar structure in COPD patients by releasing elevated amounts of inflammatory molecules [7]. The oxidant-generating systems of neutrophils produce reactive nitrogen species such as nitric oxide (NO) [8]. Inducible nitric oxide synthase (iNOS) regulated by NO and excessive production of NO are known to be associated with airway inflammation such as in COPD [91011]. A recent study showed that CS-exposed animals exhibited airway inflammation with an elevated expression of iNOS. [12]. Therefore, down-regulation of iNOS is considered an important strategy in treating CS-induced airway inflammation.
Water extract of guibi-tang (GB), a traditional herbal formula, is a mixture of 12 herbs and is known as Qui-Pi-tang in Chinese and Kihi-To in Japanese. [1314]. GB has been traditionally used to treat amnesia, fatigue, insomnia, anemia, and palpitations [15]. Studies have shown that GB has various pharmacological activities, including immunoregulatory, antioxidant, and gastroprotective effects. GB inhibits ischemic retinal angiogenesis by suppressing VEGF and PAI-1 expression and exhibits apoptotic activity by regulating mitogenactivated protein kinases (MAPKs) [1316]. However, thus far, no studies have reported the protective effects of GB against pulmonary inflammation induced by CS and lipopolysaccharide (LPS).
Therefore, we investigated the protective effects of GB on CS- and LPS-induced pulmonary inflammation in a mouse model. We evaluated the expression of several inflammatory mediators in CS- and LPS-exposed mice to explore the mechanisms of action of GB in pulmonary tissue.
Materials and Methods
Plant materials
The 12 raw herbs, Angelicae Gigantis Radix, Longan Arillus, Zizyphi Semen, Polygalae Radix, Ginseng Radix, Astragali Radix, Atractylodis Rhizoma Alba, Poria sclertum cum Pini Radix, Aucklandiae Radix, Glycyrrhizae Radix et Rhizoma, Zingiberis Rhizoma Recens, and Zizyphi Fructus were purchased from Omniherb (Yeongcheon, Korea) and from HMAX (Jecheon, Korea) in February 2008. Each raw material was verified by Professor Je-Hyun Lee, Oriental Medicine, Dongguk University (Gyeongju, Korea) and by Professor Young-Bae Seo, Oriental Medicine, Daejeon University (Daejeon, Korea). Voucher specimens (2008-KE22-1-KE22-12) have been deposited at the K-herb Research Center, Korea Institute of Oriental Medicine (KIOM, Daejeon, Korea).
Preparations of GB decoction
GB was prepared and stored at KIOM. Briefly, Angelicae Gigantis Radix (872 g), Longan Arillus (872 g), Zizyphi Semen (872 g), Polygalae Radix (872 g), Ginseng Radix (872 g), Astragali Radix (872 g), Atractylodis Rhizoma Alba (872 g), Poria sclertum cum Pini Radix (872 g), Aucklandiae Radix (436 g), Glycyrrhizae Radix et Rhizoma (262 g), Zingiberis Rhizoma Recens (1,453 g), and Zizyphi Fructus (872 g) were mixed and extracted in a 10-fold mass (100 L) of water at 100℃ for 2 h using the reflux method. The extracted water solution was freeze-dried to yield a powder (mass of extract: 2.43 kg; yield: 24.3%).
High-performance liquid chromatography (HPLC) analysis of GB sample
Liquiritin (99.6%) and glycyrrhizin (99.0%) were purchased from Wako Chemicals (Osaka, Japan). Nodakenin (98.0%) was purchased from NPC Bio Technology (Yeongi, Korea). The standard solution for reference was dissolved in methanol at a concentration of 1,000 µg/mL and stored at 4℃. The HPLC system used for the quantitative analysis of the GB sample was a Shimadzu Prominence LC-20A series instrument (LC-20A, SHIMAZU, Kyoto, Japan) coupled with a photodiode array detector. Gemini C18 (250 mm×4.6 mm; 5 µm, Phenomenex, Torrance, CA) was used for the separation of analytes and maintained at 40℃. The mobile phase system consisted of distilled water and acetonitrile, both containing 1% (v/v) acetic acid. The gradient elution program for the mobile phases was as follows: 10–70% B (0–30 min), 70–100% B (30–35 min), 100% B (for 35–40 min), 100–10% B (40–45 min), and 10% B (45–60 min). The flow rate was 1 mL/min, and the injection volume was 10 µL. For evaluation, 400 mg of freeze-dried GB extract was dissolved in 20 mL distilled water, and the solution was filtered through a 0.2-µm membrane filter (PALL Life Sciences, Ann Arbor, MI) before injecting it to the HPLC system.
Experiment animal model
Specific pathogen-free C57BL/6N mice were obtained from Samtako Co. (Osan, Korea). The animals were 6–8 weeks old and weighed 20–25 g. The mice were housed under standard conditions of temperature (22±2℃) and humidity (55±5%) and subjected to a 12-h light/dark cycle. The Institutional Animal Care and Use Committee of Chonnam National University approved the protocols for our animal study (CNU IACUC-YB-R-2016-18), and the animals were cared for in accordance with the Guidelines for Animal Experiments prescribed by Chonnam National University. The mice were divided into the following five groups: NC (non-treated mice), CS (mice exposed to CS and LPS), ROF (mice exposed to CS and LPS that received roflumilast, SML1099, Sigma-Aldrich Korea, Seoul, Korea, 10 mg/kg orally), and GB50 and GB100 (mice exposed to CS and LPS that received GB 50 and 100 mg/kg, respectively orally). The doses of GB were determined based on a previous report [13]. Roflumilast is a PDE4 inhibitor and used for COPD treatment. Roflumilast not only has bronchodilatory effects but also has protective effects against airway inflammation [1718]. Therefore, roflumilast was used as the positive control drug. For CS exposure, we used a 3R4F research cigarette, containing 9.4 mg of tar and 0.76 mg of nicotine per cigarette, and a cigarette smoke generator (Daehan Biolink, Eumseong, Republic of Korea). The mice were exposed to the cigarette fumes in a chamber (50 cm×30 cm×30 cm) for 1 h (one puff/min; 35 mL puff volume over 2 seconds; 8 cigarettes per day for 7 days). LPS (5 µg, L2630, Sigma-Aldrich Korea) was administered intranasally under mild anesthesia on day 5. Roflumilast and GB were administered by oral gavage before CS exposure on all 7 days of the experimental period.
Bronchoalveolar lavage fluid (BALF) collection and differential cell count
BALF was collected by tracheostomy of sacrificed mice 48 h after the final intranasal LPS administration. PBS (ice-cold, 0.7 mL) was infused into the lung by tracheal cannulation, and this step was repeated once (total volume 1.4 mL). To determine differential cell count, 100 µL of BALF was centrifuged on slides using a cytospin instrument (A-cyto-12, Hanil Science Industrial, Seoul, Korea). Once dried, the cells were stained with Diff-Quik® (Sysmex Corporation, Kobe, Japan) reagent. The BALF supernatant was stored at −70℃.
Pro-inflammatory cytokines measurement
Pro-inflammatory cytokine levels in BALF were measured using ELISA kits (TNF-α; DY410, IL-6; DY406, R&D System, Minneapolis, MN) according to the manufacturer's protocols. Following reaction, the plates were incubated for 10 min in the dark, and absorbance was measured at 450 nm using a microplate reader (iMark™, Bio-Rad, Hercules, CA). The amount of pro-inflammatory cytokines was calculated by measuring absorbance using a standard curve.
Western blot
Protein concentrations in homogenized (10% w/v) lung tissues were determined by using the Bradford method (#5000205, Bio-Rad). The proteins present in the tissue samples were resolved via 10% SDS-polyacrylamide gel electrophoresis, following which the electrophoresed proteins were transferred to polyvinyl difluoride (PVDF, IPVH00010, Merck, Darmstadt, Germany) membranes and incubated with a blocking solution (5% skim milk, 232100, BD Bioscience, San Jose, CA, USA) for 1 h. Appropriate primary antibodies were used to probe the membranes, and they were incubated overnight at 4℃. The primary antibodies and dilutions used were as follows: anti-β-actin (1:2,000 dilution; #4967, Cell Signaling, Danver, MA) and anti-iNOS (1:1,000 dilution; ab15323, abcam, Cambridge, UK). The membranes were washed three times using Tris-buffered saline containing Tween 20 (TBST) and incubated with 1:10,000 diluted horseradish peroxidase (HRP)-conjugated secondary antibody (Jackson Immuno Research, West Grove, PA) for 1 h at room temperature. The membranes were washed three times with TBST, and then developed using an enhanced chemiluminescence (ECL) kit (Thermo Fisher Scientific, Waltham, MA). Protein expression was calculated using densitometric band values obtained from the analysis of developed membranes in a ChemiDoc machine (ChemiDoc XRS+, Bio-Rad).
Histopathological assay
Fixed lung tissue samples were embedded in paraffin and sectioned into 4-µm-thick slices. The sections were stained with hematoxylin and eosin (H&E solution, Sigma-Aldrich) to evaluate inflammation. For the immunohistochemical assay, the paraffin-embedded lung tissues were deparaffinized, dehydrated, and washed in PBS containing 0.05% Tween 20 (PBS-T). The slides were incubated with goat serum for 20 min at room temperature to block nonspecific staining. They were then incubated for 2 h at room temperature with primary mouse antirabbit iNOS antibody (diluted 1:100, Abcam). Next, they were washed three times using PBS-T, incubated for 1 h at room temperature with a biotinylated secondary antibody, and again incubated with an avidin-biotin-peroxidase complex (Vector Laboratories, Burlingame, CA) for 1 h at room temperature. The slides were washed, incubated with diaminobenzidine (DAB, Abcam) for an additional 5 min, and examined using a microscope.
Results
HPLC analysis of GB sample
Quantitative analysis of three marker compounds in the GB sample was conducted using our optimized HPLC method. The three marker compounds, liquiritin, nodakenin, and glycyrrhizin, were eluted at 14.272, 15.205, and 30.708 min, respectively (Figure 1), and the amounts of these components in the GB sample were 1.05, 3.38, and 0.83 mg/g, respectively. These results indicate that the GB sample used in this experiment is the same as the herbal formula available on the market.
Effect of GB on inflammatory cell counts in BALF induced by CS and LPS exposure
The number of inflammatory cells was significantly higher in the CS group mice than in the NC group mice. In particular, mice from the CS group had higher neutrophil counts than mice from the NC group. Although the ROF group showed decreased inflammatory cell count in the BALF, the decrease was not significant compared to that observed in the other groups. However, mice from the GB-treated groups showed a decline in inflammatory cell counts in the BALF compared to the counts in mice from the CS group. Mice from the GB100 group, in particular, showed significantly lower neutrophil counts than mice from the CS group (Figure 2).
Effect of GB on the production of pro-inflammatory cytokines induced by CS and LPS exposure
The CS group mice showed significantly higher production of TNF-α than mice from the NC group (Figure 3A). The ROF group showed a marked decrease in the production of TNF-α compared to that of the CS group. In addition, mice from the GB-treated groups showed significantly lower TNF-α expression than mice from the CS group. Consistent with these results, IL-6 production was also significantly higher in the CS group mice than in the NC group mice (Figure 3B). By contrast, mice from the GB-treated groups showed significantly lower IL-6 levels in BALF than mice from the CS group.
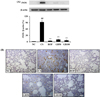
Effect of GB on iNOS expression induced by CS and LPS exposure
The results of the western blot analysis showed that the expression of iNOS was significantly higher in mice from the CS group than in mice from the NC group. The ROF group showed a significant reduction in iNOS expression compared to that in the CS group. In addition, mice from the GB-treated groups expressed significantly lower levels of iNOS than mice from the CS group (Figure 4A). Consistent with these results, iNOS expression in lung tissues was significantly higher in the CS group mice than in the lung tissues of NC group mice. By contrast, mice from the ROF and GB-treated groups had significantly lower levels of iNOS in their lung tissues than mice from the CS group (Figure 4B).
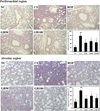
Effect of GB on inflammatory cell accumulation into lung tissue induced by CS and LPS exposure
The CS group mice showed higher inflammatory cell accumulation in the lung tissue than mice from the NC group. The ROF group showed a marked decrease in inflammatory cell infiltration into lung tissue compared to that in the CS group. In addition, mice from the GB-treated groups showed a decrease in inflammatory cell accumulation in the peribronchial and alveolar lesions compared to that observed in mice from the CS group. This suppression was most evident in the GB100 group (Figure 5).
Discussion
The incidence of COPD has been on the rise in recent years; COPD is considered a life-threatening disease due to its high morbidity and mortality [1920]. In this study, we investigated the protective effect of the herbal medicine GB against airway inflammation caused by CS and LPS. GB decreased inflammatory cell count and proinflammatory cytokine levels in the BALF of mice exposed to CS and LPS. It also effectively suppressed CS- and LPS-induced iNOS expression in the lung tissue, which was accompanied by a decrease in inflammatory cell accumulation.
CS is regarded as a critical factor in the pathogenesis of COPD, which is characterized by inflammatory responses, airway obstruction, and alteration of normal alveolar structure. [21]. Chronic exposure to CS triggers the recruitment of inflammatory cells such as neutrophils and macrophages, as CS consists of various toxic materials [22]. The infiltration of inflammatory cells into the lung tissue of COPD patients or smokers was higher than that in healthy humans [23]. Of the recruited inflammatory cells, neutrophils are involved in the development of COPD [24]. Neutrophils act as front-line defensive cells of the immune system, but excessive accumulation of neutrophils induces airway inflammation and structural alteration through the release of chemotactic mediators such as ROS, inflammatory cytokines, and tissue-damaging enzymes [2425]. Therefore, the suppression of neutrophil count is considered an important approach in controlling airway inflammation induced by CS. In this study, GB treatment effectively suppressed these elevated inflammatory cell counts in the BALF of mice exposed to CS and LPS. This was accompanied by a reduction in the accumulation of inflammatory cells in peribronchial and alveolar lesions of mice exposed to CS and LPS. These results demonstrate that GB effectively inhibited inflammatory cell accumulation induced by CS and LPS.
Proinflammatory cytokines such as TNF-α and IL-6 play a vital role in the regulation of lung inflammation induced by CS [5]. TNF-α is an important mediator of airway inflammation in COPD owing to its induction of adhesion molecules via activation of NF-κB and elevation of ROS production [2627]. In addition, IL-6 production is involved in airway inflammation and decline of lung function in COPD [28]. In COPD patients, IL-6 levels are significantly higher than the levels in healthy humans. This upregulation of IL-6 is associated with increased mortality in COPD patients [29]. In an experimental animal model, IL-6 induced not only airway inflammation but also emphysema-like airspace enlargement [30]. Thus, the extent to which TNF-α and IL-6 are downregulated serves as an important indicator of therapeutic efficacy against airway inflammation induced by CS exposure. In this study, GB treatment significantly decreased the production of TNF-α and IL-6 in BALF compared to that in CS- and LPS-exposed mice. Therefore, these results indicated that GB effectively suppressed the production of proinflammatory cytokines induced by CS and LPS exposure.
The expression of iNOS is modulated by various factors including cytokines, chemokines, and exogenous toxic materials; iNOS overexpression resulted in excessive NO production, which consequently elevated oxidative stress via NOS production and aggravated inflammatory response via activation of inflammation-related signaling [31]. iNOS should be studied in clinical trials of possible therapies because iNOS expression is closely associated with the development of COPD. It is mainly overexpressed in small airways and peripheral lung tissue of COPD patients and causes protein degradation and skeletal muscle apoptosis. [3233]. In vivo experiments with iNOS-deficient mice demonstrated inhibition of pulmonary hypertension, ventricular pressure, and vascular remodeling induced by CS exposure [34]. In this study, GB treatment significantly reduced iNOS expression induced by CS and LPS exposure. This suggests that GB effectively decreases airway inflammation resulting from CS and LPS exposure by suppressing iNOS expression.
The preventive effects of GB observed in this study were closely associated with its active components, liquiritin, nodakenin, and glycyrrhizin, which were reported to have anti-inflammatory properties in various experiments. Liquiritin effectively suppressed inflammatory responses in ultraviolet B radiation-exposed skin and decreased the levels of inflammatory mediators in cigarette smoke extract-stimulated lung epithelial cells [3536]. Nodakenin and glycyrrhizin exhibited suppressive effects on airway inflammation in a murine model of allergic asthma [3738]. Based on the properties of three components, we hypothesized that GB effectively inhibits airway inflammation in allergic asthma. Therefore, we will investigate the therapeutic effect of GB on allergic asthma in further study. In addition, we will explore the antioxidant effects of GB because its three components have antioxidant effects [3639].
Based on our results, GB effectively prevented airway inflammation induced by CS and LPS exposure. However, it is difficult to conclude that GB is effective in treating COPD because our experiment is performed with the aim of evaluating the preventive effect on airway inflammation induced by CS and LPS. Thus, additional experiment of therapeutic effect using chronic CS and LPS exposure model should be performed.


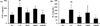
 XML Download
XML Download