

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Increased levels of apolipoprotein CIII (apoCIII) in sera from patients with type-1 diabetes mellitus (T1D) have been shown to promote Ca2+-induced β-cell apoptosis [1]. Although the exact molecular mechanisms are not known, we have demonstrated that apoCIII hyperactivates the voltage-gated L-type Ca2+-channel through scavenger-receptor class BI (SR-BI)/β1 integrin-dependent coactivation of PKA and Src [2].
The diabetes-prone BioBreeding (DP-BB) rat colony was established in the 1970s in Ottawa, Canada from outbred Wistar rats that spontaneously developed a human-like autoimmune T1D. Both sexes of DP-BB rats develop pancreatic insulitis, morphologically similar to that observed in human T1D, with a selective destruction of the insulin-secreting β-cells. In our breeding colony DP-BB rats become diabetic when they are around 60 days old.
The pre-diabetic rats have, as human T1D patients, increased serum concentration of apoCIII [3]. We have decreased the endogenous levels of apoCIII by antisense treatment between the age of 12 to 40 days, when they are still in the pre-diabetic phase, and that significantly prolonged the time to onset of diabetes [3].
ApoCIII is an 8.8 kDa polypeptide mainly synthesized in the liver. There are several pathways so far known to be involved in the regulation of apoCIII gene expression. Interestingly, the gene for apoCIII is inversely regulated by insulin, i.e. insulin inhibits transcription while insulin resistance, seen in many type-2 diabetes (T2D) patients, leads to an increase in apoCIII [45]. Other regulators are peroxisome proliferator-activated receptors (PPARs) that reduce apoCIII gene expression [6].
There are several studies where the beneficial effects of polyphenols in wine have been discussed [78] including data showing a decrease in liver mRNA levels of apoCIII [9]. ApoCIII is pro-inflammatory and polyphenol-rich grape products have been suggested to increase the expression or activation of PPARs that antagonize inflammatory transcription factors [8]. There are data showing that streptozotocin-induced diabetic rats have a reduced anti-oxidant capacity, compared to normal rats, which could be restored by administration of polyphenol enriched Chardonnay white wine. The effect was alcohol-independent [10].
Fibrates are used in the treatment of hyperlipidemia and have, among several other actions, been reported to have anti-inflammatory effects and to reduce apoCIII through the activation of PPARα [1112]. There are emerging data that PPARα is also of importance for diabetes induced microvascular complications like retinopathy [1112].
The aim of this study, based on the data that the proinflammatory apoCIII is increased in diabetes [1314], was to investigate if treatments with polyphenol-rich red wine or fenofibrate have any influence on the onset of diabetes in our animal model for T1D, the BB-rat. These animals develop diabetes within a very narrow time window which gives unique opportunities to study the impact of pre-diabetic interventions.
Materials and Methods
Animals
Diabetes Prone Bio Breeding (DP-BB) rats were obtained from our breeding colony. The incidence of diabetes among our rats is 100% with a mean age of onset at 60 days. Diabetes onset is defined by a glucose level of 270 mg/dL or above. The animals were housed under SPF (Specific Pathogen Free) conditions in a temperature- and humidity-controlled room with 12 hours light: dark cycles. They were fed the R36 diet and water ad libitum. All experiments were carried out according to the local Animal Experiment Ethics Committees.
Red wine treatment
Rats were administered 3.2 mL/kg of red wine (equal to one glass of wine/day) by oral gavage. The treatment was given for one month and started when the rats were 30 days old. Two different kinds of wine were used; Sonovino Primitivo, Italy consisting of 90% Primitivo grapes and 10% Negroamaro containing medium amounts of polyphenols and Reserve du Vieux Noir, Malbec, France made of Malbec grapes containing high levels of polyphenols.
To rule out an effect of alcohol rats received wine where the alcohol had been removed by evaporation, subjecting the wine to 50℃ for 30 min. Control rats were given water by oral gavage.
Fenofibrate treatment
Rats were administered a daily dose of 100 mg/kg Fenofibrate (Lipanthyl, Abbott, France) dissolved in 1% Methyl cellulose solution (MTC) by oral gavage. The treatment was given for one month and started when the rats were 30 days old. Controls were given the vehicle.
In both treatment groups rats were weighed each day until onset of diabetes. If a rat developed diabetes before the age of 60 days treatment was terminated. Blood samples were taken before and after 30 days of treatment, and at the debut of diabetes.
RNA isolation and quantitative RT-PCR
Total RNA was isolated using the RNeasy Mini Kit according to the manufactures protocol (Qiagen, Germantown, MD, USA). Total RNA was reverse transcribed at 37℃ using the High Capacity cDNA Reverse Transcription kit (Life technologies, Carlsbad, CA, USA). The expression of all genes was measured by real-time quantitative PCR with Maxima SYBR Green qPCR Master Mix with ROX (Thermo scientific, Waltham, MA, USA) on a QuantStudio 5 instrument (Life technologies). β-Actin was used as an endogenous control. Primer sequences will be provided upon request.
Statistical analysis
Statistical analyzes were performed with GraphPad Prism. For individual experiment, the number of animals used (n) is included in each figure legend in parenthesis. All results are expressed as mean±SEM. A student's t-test or one-way ANOVA (Tukey's post-hoc) were used when appropriate. P values <0.05 were considered statistically significant, however, this level was not reached in any of the experiments.
Results
The BB-rat animal model for T1D is suitable for intervention studies as the onset of diabetes appears within a narrow time window around 60 days of age and all DP-BB rats in our colony develop the disease. We have previously shown that lowering apoCIII by antisense treatment during the prediabetic phase prolongs the time to onset of disease [3]. In this study we wanted to examine whether red wine containing polyphenols or the lipid-lowering drug, fenofibrate, both reported to decrease apoCIII, have any effect on the debut of diabetes.
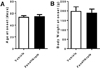
In the wine experiments the rats received a daily dose of red wine equivalent to one standard glass. The wine, and the water to the control rats, were given by oral gavage to ensure that they all got the same dose. Two brands of wine with different contents of polyphenols were tested. The animals were treated for one month, but we could not see any effect on the onset of disease by the different wines (Figure 1A). Body weight did not differ significantly, although there seemed to be a tendency to lower weight in both wine groups (Figure 1B).
In the second experiment fenofibrate, that is mainly used to treat hypertriglyceridemia, was given once per day and, as with the wine, it was administered by oral gavage. Neither did we see any effects of this treatment. The age at onset of T1D and body weight were similar in rats treated with fenofibrate as to those receiving the vehicle (Figure 2A, B).
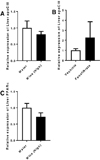
Since our hypothesis was that polyphenols and fenofibrate, by reducing the levels of the pro-inflammatory lipoprotein apoCIII will affect the onset of diabetes, the expression levels of apoCIII were analyzed in liver samples, as liver is the main source of apoCIII. None of the treatments lowered the expression of the lipoprotein (Figure 3A, B). As polyphenols have been reported to up-regulate the expression of PPARα, while fenofibrate activates the receptor, the expression of PPARα was analysed in livers from rats that had been given wine with high concentration of polyphenols, but the levels were similar to those given water (Figure 3C).
Discussion
ApoCIII is well-known within the field of cardiovascular research, where increased levels have been demonstrated to be pro-inflammatory resulting in atherosclerosis and higher risk for cardiovascular diseases (CVD) [1516]. It has convincingly been shown in both human and animal studies that there is a connection between apoCIII, hypertriglyceridemia and CVD [1718].
ApoCIII has not only been related to CVD, but also to diabetes. It has been demonstrated that haplotypes in the apoCIII gene leading to augmented levels of apoCIII are associated with an increased susceptibility to T1D [19]. Furthermore, apoCIII gene variants with increased levels of apoCIII are associated with the development of non-alcoholic fatty liver disease (NAFLD), hepatic insulin resistance and T2D [2021]. On the contrary, there are humans with mutations in the apoCIII gene leading to life-long reduced levels of apoCIII. These individuals are healthier with favorable pattern of lipoproteins, increased insulin sensitivity, lower incidence of hypertension and they live longer [222324]. Hence, there are a multitude of beneficial health effects lowering apoCIII.
There are several factors involved in development of T1D. We have previously demonstrated that sera from patients with T1D contain increased levels of apoCIII and that this promotes Ca2+-induced β-cell apoptosis that can be prevented by reducing apoCIII [13]. That increased levels of apoCIII are indeed of importance for β-cell function and survival was demonstrated in vivo in the T1D BB rat animal model. When apoCIII was lowered by antisense during a phase of the pre-diabetic period the time to onset of diabetes was prolonged by comparable eight human years [3].
ApoCIII has also been shown to be increased in patients with type-2 diabetes (T2D) [2526] and it is known that insulin resistance, as well as deficiency of insulin, up-regulates the apoCIII gene [45].
During many years there have been discussions whether a moderate consumption of red wine is beneficial. It has for example been shown that red wine rich in polyphenols with and without alcohol improves insulin sensitivity and activates anti-inflammatory and anti-oxidant processes with positive effects on CVD [2728]. Furthermore, the use of lipid lowering substances has been reported not only to reduce the risk of CVD by lowering lipids, but also to have anti-inflammatory effects [29]. Both polyphenols and fibrate have been reported, among a lot of other effects, to lower apoCIII. Since it would be desirable to use an alternative to antisense to decrease apoCIII, we decided to investigate if the onset of T1D in our BB rat animal model could be affected by treating with polyphenol containing wine or fibrate.
We tested two red wines (without alcohol) with different content of polyphenols and fenofibrate, but neither of the treatments had any influence on the debut of T1D. When measuring the expression levels of apoCIII there was no difference which is in line with the observation that the onset of T1D was unaffected.
Rats treated with wine had a tendency to a lower body weight. This has been observed in other studies that a moderate consumption of wine decreases weight and even partially prevents high-fat diet induced weight gain [3031].
In a study in Sprague-Dawley rats, 2 and 24- months old, treated with fenofibrate for two weeks, an age-related reduced expression of PPARα was seen, but no effect by fenofibrate although there was a decrease in triglycerides showing that the drug was absorbed [32]. However, apoCIII mRNA levels were reduced. The authors suggested that this could reflect a functional activation of PPARα despite no change in expression.
There could be several explanations why there is a discrepancy to our results. Important differences are the age of the animals, the treatment time and the use of a different strain, the diabetes-prone BB rat is on a Wistar background. Furthermore, the Sprague-Dawley rats were not in a pre-diabetic stage where many different pathways can be affected, prior to the onset of disease, influencing the responses.
In conclusion, we demonstrate that neither red wine nor fenofibrate lowered apoCIII or affected the onset of diabetes in the BB rat T1D animal model. However, it should be emphasized that this does not exclude that other species may show a different response to these substances. Since apoCIII is regulated by several different pathways it might be necessary to lower it either at the transcription level with antisense, as we have shown before [3], or at the protein level by inhibiting the function using an antibody, nanobody or aptamer against apoCIII.

 XML Download
XML Download