

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Benign prostate hyperplasia (BPH), a nonmalignant enlargement of the prostate gland, is a common disease in elderly men [1]. It was reported that 50% of men over the age of 50 show histological evidence of BPH, which is characterized by significant proliferation of the epithelial cells within the prostate gland [2]. Although its pathogenesis is a complex multi-component process, it is well known that dihydrotestosterone (DHT) plays an important role in the progression of BPH [3]. DHT is formed upon the conversion of testosterone by 5α-reductase, which leads to an increased level of DHT in the prostate. Excessive DHT stimulates prostate tissue proliferation via binding with α-adrenergic receptors, which causes enlargement of the prostate gland [4]. Therefore, a useful strategy for the treatment of BPH may be the reduction of the conversion of testosterone into DHT through the inhibition of 5α-reductase. In the pharmacological approach to the treatment of BPH, 5α-reductase inhibitors such as finasteride and dutasteride are most commonly used in conventional medicine [5]. However, 5α-reductase inhibitors may have serious side-effects, such as orthostatic hypotension, decreased libido and ejaculation, or erectile dysfunction [6]. Owing to the small number of adverse effects of natural products, they have been considered as alternative treatments for BPH [7].
Pycnogenol® (PYC), a standardized French maritime pine bark extract, is composed of procyanidins and phenolic acids [8]. It is a well-known antioxidant agent and is used for the treatment of various medical conditions caused by oxidative stress, such as nephrotoxicity [9], cardiotoxicity [10], hepatotoxicity [11], and ototoxicity [12]. In particular, PYC was reported to have the potential to inhibit the proliferation of human cancer cell lines through the induction of apoptosis [13]. Despite these antiproliferative effects, there are no published studies on the protective effects of PYC against BPH.
Therefore, in this study, we investigated the protective effects of PYC in a rat model of testosterone propionate (TP)-induced BPH through the measurement of changes in the weight of the prostate, serum DHT concentration, histology, and the expression of proliferative markers.
Materials and Methods
Animals and environmental conditions
Twelve-week-old male Sprague-Dawley rats were obtained from a specific-pathogen-free colony at Samako Co. (Osan, Korea) and were used after 1 week of quarantine and acclimatization. Two rats per cage were housed in stainless wire mesh cages in a room maintained at constant conditions (temperature, 23±3℃; relative humidity, 50%±10%; artificial lighting, 08:00–20:00; air changes per hour, 13–18). The rats were provided with tap water that had been sterilized by ultraviolet irradiation and commercial rodent chow (Samyang Feed, Wonga, Korea) ad libitum. The Institutional Animal Care and Use Committee of Chonnam National University approved the protocols for the animal study (CNU IACUC-YB-R-2016-38), and the animals were cared for in accordance with the Guidelines for Animal Experiments of Chonnam National University.
Experimental procedure
After acclimatization, the rats were divided into five groups: (A) a normal control group administered phosphate-buffered saline (PBS, p.o.) with corn oil (s.c.); (B) a BPH group administered PBS (p.o.) and testosterone propionate (TP, 3 mg/kg, s.c.); (C) a positive drug control group administered finasteride (10 mg/kg, p.o.) with TP (s.c.); and (D and E) PYC-treated groups that received PYC at 20 or 40 mg/kg (p.o.), respectively, with TP (s.c.). TP was purchased from Tokyo Chemical Ins. Co. (Tokyo, Japan) and finasteride, a 5α-reductase inhibitor used as a positive control anti-BPH drug, was purchased from Sigma-Aldrich (St Louis, MO, USA). The effective dose of finasteride for the treatment of BPH was determined from a previous study [14]. All materials were administered to animals once daily for 4 weeks, and body weight was measured weekly. The application volumes were 5 mL/kg for p.o. administration (PBS, finasteride, and PYC) and 3 mL/kg for s.c. injection (corn oil and TP) and were calculated in advance based on the most recently recorded body weight of each animal.
Blood and tissue samples
After the final treatment, all animals were fasted overnight and euthanized by the intraperitoneal injection of Zoletil 50® (Virbac Laboratories, Carrots, France) at 10 mg/kg body weight. Blood samples were drawn from the caudal vena cava, and the serum was separated by centrifugation and stored at −80℃ until used for hormone assays. The prostate was removed immediately and weighed. The relative prostate weight was calculated as the ratio of prostate weight to body weight. The ventral lobe of the prostate was divided in half. One half was fixed using 10% neutral-buffered formalin and embedded in paraffin for histopathological examination and the other was stored at −80℃ for western blot analysis.
Measurement of DHT concentration in serum and the prostate
The concentration of DHT in the serum and the prostate was determined by using an enzyme-linked immunosorbent assay (ELISA) kit in accordance with the manufacturer's instructions (ALPCO Diagnostics, Salem, NH, USA). The absorbance was measured at 450 nm by using a microplate ELISA reader (Bio-Rad Laboratories, Hercules, CA, USA). The values are expressed in milliliters for serum and in milligrams for the prostate.
Histopathological examination
The prostate tissue was fixed in 10% (v/v) neutral buffered formalin, embedded in paraffin, sectioned to 4-µm thickness, and stained with hematoxylin and eosin (H&E) solution (BBC Biochemical, Mount Vernon, WA, USA) to assess the morphological changes in the prostate. All observations were performed manually in a completely blinded manner by using a light microscope (Leica DM LB2; Leica, Wentzler, Germany) with ×10 and ×20 objective lenses and a ×100 oil immersion lens.
Immunohistochemical staining of Ki-67
To determine Ki-67 expression, we performed immunohistochemical analysis by using Vector stain ABC IHC kit (Vector Laboratories, Burlingame, CA, USA) in accordance with the manufacturer's protocol. The antimouse Ki-67 antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) was used at a 1:200 dilution. Each slide was examined manually in a completely blinded manner by using light microscopy (Leica) with ×10 and ×20 objective lenses and a ×100 oil immersion lens. Quantitative analysis of Ki-67 expression was performed by using an image analyzer (IMT i-Solution software, Vancouver, BC, Canada) on 10 randomly selected nonoverlapping areas per slide were captured by using a digital camera (IMTcamCCD5; IMT Inc., Daejeon, Korea).
Western blot analysis for proliferating cell nuclear antigen (PCNA)
The prostate tissue was homogenized by using a homogenizer in a tissue lysis/extraction reagent (1/10 [w/v]; Sigma-Aldrich) containing a protease inhibitor (Roche, Basel, Switzerland). The protein concentration for each sample was determined by using the Bradford reagent (Bio-Rad Laboratories). Western blotting was performed as described previously [14] by using the following antibodies: anti-PCNA (1:1,000; Abcam, Cambridge, UK); anti-β-actin (Cell Signaling Technology, Beverly, MA, USA). To determine protein expression, the density of each band was quantified by using ChemiDoc (Bio-Rad Laboratories).
Statistical analyses
The data were expressed as the mean ± SD. All statistical comparisons were made by using one-way analysis of variance followed by Tukey's multiple comparison test. Statistical significance was computed by using GraphPad InStat v. 3.0 (GraphPad Software, La Jolla, CA, USA). The differences were considered statistically significant for values of P<0.05 or P<0.01.
Go to : 
Results
Effect of PYC on relative prostate weight
As shown in Figure 1, the relative prostate weight from rats in the BPH group was significantly higher than that in the normal control group. In contrast, compared with the BPH group, the rats in the PYC-treated groups had significantly decreased relative prostate weights, which were similar to those in the finasteride-treated group. In particular, 40 mg/kg PYC treatment resulted in a significant reduction in the relative prostate weight compared with the BPH group.
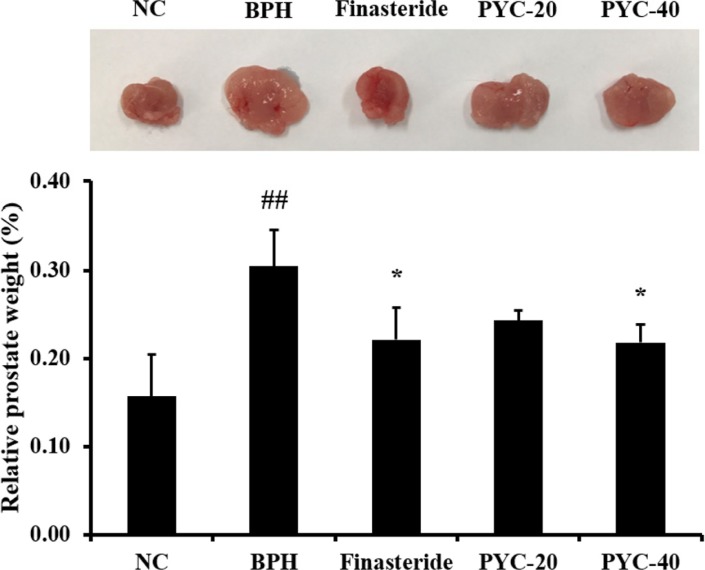 | Figure 1The effects of PYC on relative prostate weights in TP-treated rats. NC, corn oil injection (s.c.)+PBS (p.o.); BPH, TP (s.c.)+PBS (p.o.); Finasteride, TP (s.c.)+finasteride (10 mg/kg, p.o.); PYC-20, TP (s.c.)+PYC (20 mg/kg, p.o.); PYC-40, TP (s.c.)+PYC (40 mg/kg, p.o.). ##P<0.01 vs. the NC. *P<0.05 vs. the BPH. The values are expressed as the mean±SD (n=6).
|
Effect of PYC on DHT concentration in the serum and prostate
The serum DHT concentration was markedly higher in the BPH group than in the normal control group. However, serum DHT concentration in the finasteride- and PYC-treated groups was significantly lower than that in the BPH group (Figure 2A). Similar to serum DHT, prostate DHT concentration was significantly higher in the BPH group, but markedly lower in the PYC-treated groups (Figure 2B).
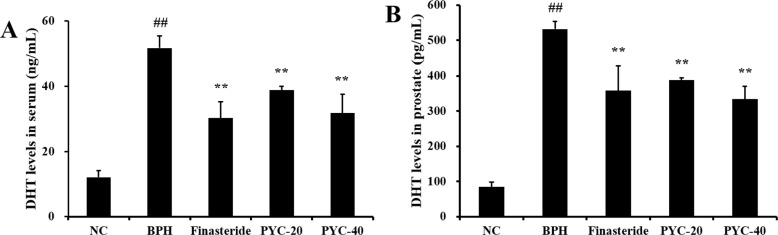 | Figure 2The effects of PYC on (A) DHT concentration in the serum and (B) in the prostate. NC, corn oil injection (s.c.)+PBS (p.o.); BPH, TP (s.c.)+PBS (p.o.); Finasteride, TP (s.c.)+finasteride (10 mg/kg, p.o.); PYC-20, TP (s.c.)+PYC (20 mg/kg, p.o.); PYC-40, TP (s.c.)+PYC (40 mg/kg, p.o.). ##P<0.01 vs. the NC. **P<0.01 vs. the BPH. The values are expressed as the mean±SD (n=6).
|
Effect of PYC on prostatic epithelial hyperplasia
Prostate epithelial cell layers were thicker in the TP-induced BPH group than in the normal control group. The finasteride-treated group showed mild epithelial hyperplasia compared with the BPH group. Similar to the finasteride-treated group, the epithelial cell layers of the prostate were thinner in the PYC-treated groups than in the BPH group (Figure 3).
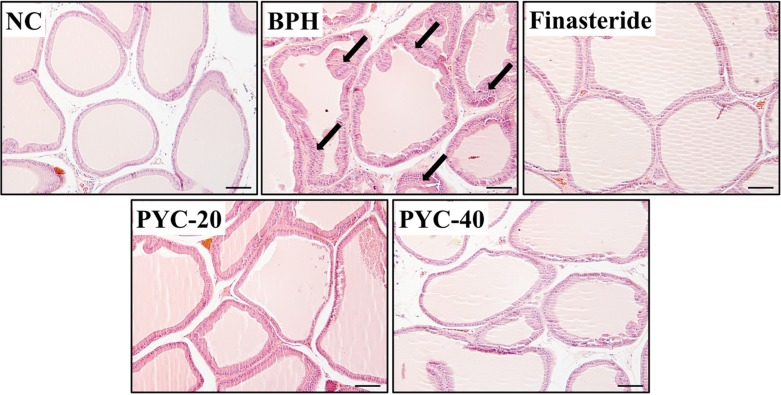 | Figure 3The effects of PYC on prostate hyperplasia. The histological examination of prostate tissue was performed by using H&E staining. NC, corn oil injection (s.c.)+PBS (p.o.); BPH, TP (s.c.)+PBS (p.o.); Finasteride, TP (s.c.)+finasteride (10 mg/kg, p.o.); PYC-20, TP (s.c.)+PYC (20 mg/kg, p.o.); PYC-40, TP (s.c.)+PYC (40 mg/kg, p.o.). Arrows indicate the area of prostate hyperplasia. Scale bar=50 µm (×200).
|
Effect of PYC on expression of Ki-67
To evaluate the expression of Ki-67, we performed IHC in the prostate tissues of each group. The protein expression of Ki-67 in the prostate tissue was higher in the BPH group than in the normal control group. However, finasteride- and PYC-treated groups obviously decreased Ki-67 expression in prostate epithelial cells in comparison with the BPH group (Figure 4).
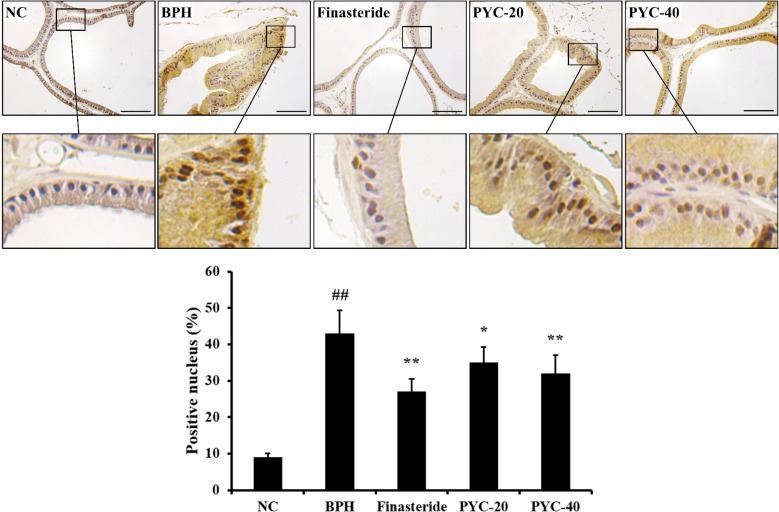 | Figure 4The effects of PYC on the protein expression of Ki-67 in the prostate. The histological examination of prostate tissue was performed by using immunohistochemistry. NC, corn oil injection (s.c.)+PBS (p.o.); BPH, TP (s.c.)+PBS (p.o.); Finasteride, TP (s.c.)+finasteride (10 mg/kg, p.o.); PYC-20, TP (s.c.)+PYC (20 mg/kg, p.o.); PYC-40, TP (s.c.)+PYC (40 mg/kg, p.o.). ##P<0.01 vs. the NC. *, **P<0.05 and P<0.01 vs. the BPH, respectively. The values are expressed as the mean±SD (n=6). Scale bar=50 µm (×400).
|
Effect of PYC on expression of PCNA protein in the prostate
Protein expression levels of PCNA, an indication of proliferation, was markedly higher in the BPH group than in the normal control group (Figure 5). However, finasteride- and PYC-treatment resulted in significantly lower expression of PCNA than those in the BPH group.
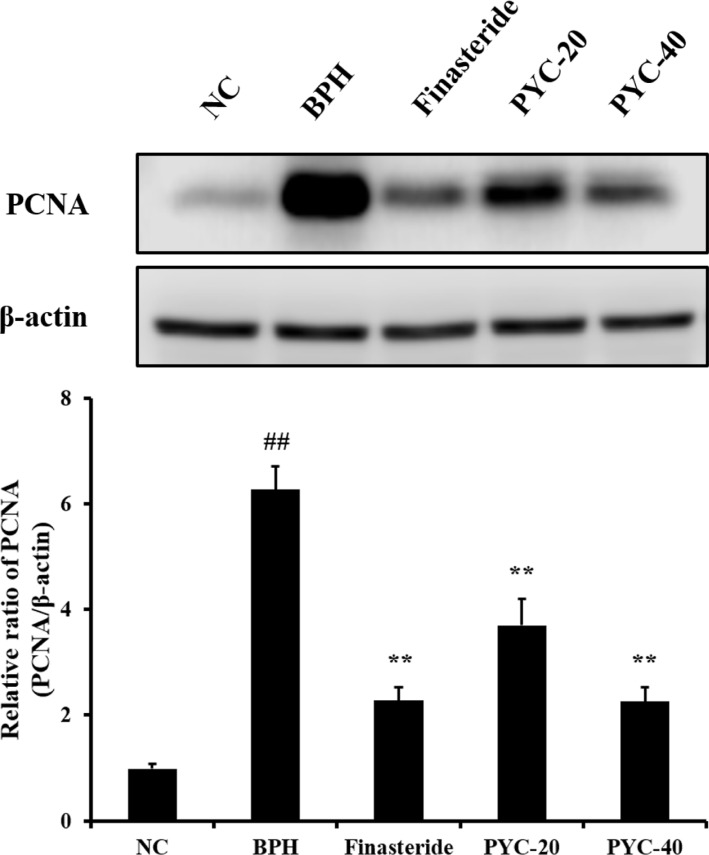 | Figure 5The effects of PYC on the protein expression of PCNA. Images of the gel and relative ratio of PCNA expression (ratio of PCNA to β-actin). NC, corn oil injection (s.c.)+PBS (p.o.); BPH, TP (s.c.)+PBS (p.o.); Finasteride, TP (s.c.)+finasteride (10 mg/kg, p.o.); PYC-20, TP (s.c.)+PYC (20 mg/kg, p.o.); PYC-40, TP (s.c.)+PYC (40 mg/kg, p.o.). ##P<0.01 vs. the NC. **P<0.01 vs. the BPH. The values are expressed as the mean±SD (n=6).
|
Go to :
Discussion
BPH, the most common benign reproductive disease in males, has become an increasingly important disease in the aging society [15]. In the present study, we evaluated the inhibitory effects of PYC on the development of BPH by using a rat model of TP-induced BPH. Animals with BPH showed increases in the prostate relative weight and DHT concentration in the serum and the prostate. The histopathological examinations revealed prostate epithelial cell hyperplasia and the overexpression of Ki-67 in addition to elevated PCNA expression. In contrast, PYC-treated animals showed a decrease in relative weight of the prostate and DHT concentration in the serum and prostate, which was associated with mild prostate epithelial cell hyperplasia and lower expression of Ki-67 and PCNA.
Rats with BPH had significantly higher relative prostate weight than the normal control group. However, administration of PYC resulted in a significant reduction of relative weight of the prostate compared with the BPH animals. BPH is characterized by stromal and epithelial cell hyperplasia, which results in prostate enlargement [4]. Many studies have shown that animals with BPH had significantly higher prostate weights than in normal control animals [1617]. In clinical trials, patients with BPH or other related diseases, such as lower urinary tract symptoms, resulted in an enlargement of the prostate gland compared with healthy subjects [18]. For this reason, an increase in prostate weight is used as an indication of BPH. These results were consistent with the histopathological findings from the prostate examination. BPH animals exhibited epithelial cell hyperplasia, whereas animals treated with PYC showed mild epithelial cell hyperplasia. According to Harati et al. [19], PYC had the potential to induce apoptosis and alter gene expression in fibrosarcoma cells. In particular, PYC inhibited cell proliferation in a dose- and time-dependent manner in human promyeloid leukemia cells [13]. In addition, the administration of PYC resulted in the reduction of protein expression of PCNA and Ki-67 induced by TP. Ki-67 and PCNA proteins are widely used to assess cellular proliferation [20]. In particular, previous studies showed that both proteins were correlated with BPH [2122]. Therefore, the reduced expression of these proteins strongly supported the antiproliferative effects of PYC. Collectively, it was thought that the role of PYC in the inhibition of prostate weight gain by TP was mediated by the inhibition of differentiation that leads to prostate epithelial cell hyperplasia.
DHT is a metabolite of testosterone produced by 5α-reductase [3]. In the various causes of BPH pathogenesis, DHT is known as a hormone that plays a particularly crucial role in the development of BPH. It binds to nuclear androgen receptors and signals the transcription of growth factors that are mitogenic for epithelial and stromal cells, which results in prostate stromal and epithelial cell hyperplasia [4]. As DHT has a higher affinity for the androgen receptor than testosterone and dissociates more slowly at the androgen receptor, the inhibition of DHT production is considered to be an important strategy to prevent the deterioration of BPH [23]. For this reason, many researchers have reduced DHT levels through the inhibition of 5α-reductase and, for a clinical approach, 5α-reductase inhibitors, such as finasteride, are commonly used as treatments of BPH [5]. In the present study, BPH animals showed increased DHT concentration in the serum and the prostate, whereas finasteride and PYC treatment reduced these increases induced by TP. These results indicated that PYC inhibited the development of BPH in rats and that these effects were closely related to the reduction in DHT concentration in animals with BPH.
In conclusion, the oral administration of PYC in a rat model of BPH significantly decreased the weight of the prostate and prostatic hyperplasia, which resulted from reduced DHT concentrations in the serum and prostate. These results were supported by inhibition of the expression of proliferative markers, including PCNA and KI-67. Our findings thus strongly suggested that PYC may be a useful agent for the treatment of BPH.
Go to :
 XML Download
XML Download