

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Renal ischemia reperfusion (IR) injury is an important clinical condition and causes significant morbidity and mortality during clinical procedures, such as kidney transplantation, partial nephrectomy, and hemorrhagic shock. Among various pathophysiological mechanisms of IR injury, oxidative stress plays a key role in inducing cellular and tissue damage. Excessive production of reactive oxygen species (ROS) and suppression of anti-oxidant defense system have been found to contribute to IR insult in many organs. Furthermore, ROS can activate the mitogen-activated protein kinase (MAPK) pathway, which plays a pivotal role in signal transduction during IR injury.12 Three distinct subgroups of MAPK, including extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38, are involved in cell recovery, survival, and death during IR injury.34
Gaseous transmitters have been implicated in the regulation of multiple pathophysiological functions in mammalian tissues.5 Along with nitric oxide (NO) and carbon monoxide (CO), hydrogen sulfide (H2S) has been characterized as a gaseous bioactive substance in various body systems. Recently, H2S has emerged as an endogenous molecule with important physiological roles. The cytoprotective or beneficial effects of the H2S donor sodium hydrosulfide (NaHS) against myocardial infarction, gastric ulcer, hypertension, and IR injury have been observed in several animal models.678
H2S is synthesized from cysteine by cystathionine γ-lyase (CGL), cystathionine β-synthase (CBS), and 3-mercaptopyruvate sulfur transferase (3-MST). Although all of these enzymes are required for H2S production, CGL and CBS are the major enzymes.9 These two enzymes are expressed in a variety of tissues and organs, including the kidney, liver, central nervous system, and vascular system.1011 The kidney is the important organ for endogenous H2S production by CGL and CBS.12 Inhibition of CGL and CBS, using D,L-propargylglycine (PAG) and amino-oxyacetic acid (AOAA), respectively, is a common strategy for investigating the effect of endogenous H2S on a disease.
In this study, we investigated whether exogenous H2S protects against renal IR injury, and the effects of inhibition of CGL or CBS were assessed to determine the relative contribution of both enzymes on H2S production in renal IR injury. Moreover, we intended to elucidate the underlying mechanism of action of H2S in the kidneys through the MAPK family.
MATERIALS AND METHODS
Animals
All experiments were performed on male Sprague-Dawley rats (280–300 g), which were obtained from Central Laboratories (Seoul, Korea). All protocols were approved by the Institutional Animal Care and Use Committee at Kyungpook National University (2015-0029), Daegu, Korea.
Experimental protocols
The rats were randomly assigned to one of five groups (n=5 per group). Group 1 was the sham group (sham); the rats underwent a sham operation without the renal IR procedure. Group 2 was the renal IR control group (IR control); the rats underwent the renal IR procedure. Group 3 was the H2S donor administration group (NaHS); the rats received 100 µmol/kg NaHS by intraperitoneal (i.p.) injection 30 min before the renal IR procedure. Group 4 was the AOAA administration group (AOAA); the rats received 10 mg/kg AOAA by i.p. injection 30 min before the renal IR procedure. Group 5 was the PAG administration group (PAG); the rats received 50 mg/kg of PAG by i.p. injection 30 min before the renal IR procedure. The dose of each drug was selected based on previous studies.131415 Rats were anesthetized with ketamine (60 mg/kg, i.p.) and xylene (10 mg/kg, i.p.) prior to the renal IR procedure. To perform the renal IR procedure, first, the right nephrectomy was carried out. Next, the left kidney vasculature was clamped for 30 min using a nontraumatic vascular clamp. The clamp was then removed to allow reperfusion. After reperfusion for 24 h, blood samples were collected from the heart and centrifuged at 3000 rpm for 15 min for analysis of blood urea nitrogen (BUN) and creatinine (Cr) levels. A portion of the harvested renal tissues was stored in liquid nitrogen at −80℃ for the analysis of malondialdehyde (MDA), superoxide dismutase (SOD), and MAPK family protein expression. The remaining tissues were processed for histopathological examination.
Biochemical analysis
To assess renal functions, serum BUN and Cr levels were analyzed using the colorimetric assay according to the manufacturer's protocols (Asan Pharmacy, Seoul, Korea). MDA level in the renal tissue was quantified using thiobarbituric acid reactive substances, and the absorbance of the reaction mixture was measured at 535 nm. The levels were expressed as nmol/g of tissue, according to a standard curve.16 SOD activity in the renal tissue was assessed using the pyrogallol autoxidation method, and the absorbance was measured at 420 nm.17 SOD activity was calculated as U/mg of protein.
Western blot analysis
Renal tissues were homogenized using a glass homogenizer (Kontes Glass Co, Vineland, NJ, USA) in RIPA buffer (Thermo Scientific, Waltham, MA, USA) containing protease and a phosphatase inhibitor cocktail (Thermo Scientific). The samples were centrifuged at 12500 rpm for 20 min at 4℃ followed by separation of the supernatant. Total protein was quantified using the Bradford assay (Thermo Scientific). The samples were mixed with loading buffer solution [60 mM Tris-HCl, 25% glycerol, 2% sodium dodecyl sulfate (SDS), 14.4 mM 2-mercaptoethanol, and 0.1% bromophenol blue], separated on a 10% SDS polyacrylamide gel, and transferred to a nitrocellulose membrane (Whatman GmbH, Dassel, Germany). The membrane was blocked with 1% bovine serum albumin in Tris-buffered saline containing 0.1% Tween-20 (TBS-T) for 1 h at room temperature. Following washes, the blots were incubated overnight at 4℃ with antibodies against p-JNK, p-ERK, p-p38 (diluted 1:1000; Santa Cruz, CA, USA), and β-actin (diluted 1:5000; Cell Signaling, Beverly, MA, USA). Membranes were then incubated with horseradish peroxidase-conjugated secondary antibody (diluted 1:1000; Cell Signaling Technology), developed using the enhanced chemiluminescence substrate kit (Advansta, Menlo Park, CA, USA), and exposed onto Agfa medical X-ray film (Agfa-Gevaert Group, Mortsel, Belgium). The signal intensity was quantified using the NIH Image J 1.47v software (National Institutes of Health, Bethesda, MD, USA).
Histopathological analysis
Renal tissues were immediately perfused in 10% phosphate-buffered formalin, embedded in paraffin, sectioned to a 4-micrometer thickness, and stained with periodic acid Schiff (PAS). The sections were evaluated under a light microscope at 200× magnification, and five fields per slide were scored by tubular cell form, luminal state, and cast formation in a blind fashion. They were then graded on a scale of 0 to 3 (0: normal; 1: minimal; 2: moderate; 3: severe) by a renal pathologist in an unbiased fashion.18
Apoptosis analysis
The terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay was performed as an index of cell injury using a detection kit (Fluorescein, Roche, Mannheim, Germany) according to the manufacturer's instructions. In brief, kidney sections were incubated with TUNEL reagent mixture, followed by washing with phosphate-buffered saline. 4,6-diamidino-2-phenylindole (DAPI) was used for nuclei staining. Images of TUNEL staining were collected from the outer medulla of kidneys using a microscope at 400× magnification (Leica DM2500). TUNEL-positive cells were counted in an unbiased fashion in five random fields per kidney sample under a light microscope with 400× magnification.
Statistical analysis
Data analysis was performed using SPSS software (Version 18.0 for Windows; SPSS Inc., Chicago, IL, USA). Data are expressed as a mean±SEM. The Mann-Whitney U test was used to assess the appropriateness of renal IR injury between the sham group and the IR control group. After performing the Kruskal-Wallis test to detect differences among the four groups, we applied the Mann-Whitney U post hoc test, each with Bonferroni correction, for the three pairwise comparison of the renal IR group with the other treatment groups. Differences between the groups were considered significant at p<0.05.
RESULTS
Changes in renal function
Induction of renal ischemia (30 min) followed by reperfusion (24 h) resulted in a significant elevation of serum Cr and BUN levels, compared to those in the sham group (p=0.008 for both). Administration of NaHS significantly attenuated this renal IR-induced elevation of serum Cr and BUN levels (p=0.024 for both). In the AOAA and PAG groups, serum Cr and BUN levels did not show any significant difference, compared to those in the IR control group (Fig. 1).
Changes in renal oxidative stress
MDA levels in the IR control group were significantly increased, compared to those in the sham group (p=0.008). Administration of NaHS significantly attenuated the elevation of MDA levels caused by renal IR injury (p=0.024). SOD levels in the IR control group were significantly lower than those in the sham group (p=0.008). Administration of NaHS significantly preserved SOD levels, compared to those in the IR control group (p=0.024). SOD levels were also preserved in the AOAA and PAG groups, compared to those in the IR control group (p=0.024 for both) (Fig. 2).
Changes in renal MAPKs
Renal IR promoted phosphorylation of ERK, JNK, and p38 (p=0.008 for all). NaHS significantly inhibited renal IR-induced activation of MAPK pathways (p=0.024 for all). Phosphorylation of ERK was decreased in the PAG group, compared to that in the IR control group (p=0.024). Rats in the AOAA and PAG groups showed increased levels of phosphorylated ERK, JNK, and p38, similar to those observed in the renal IR control group (Fig. 3).
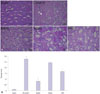
Histopathological evaluation
Analysis of tissue sections from the kidneys by PAS staining demonstrated a significant tissue damage in the renal IR control group (p=0.008). This was manifested by tubular dilatation, luminal congestion, and tubular cell atrophy. In the NaHS group, kidney structures appeared better, with mild tubular dilatation and luminal congestion (p=0.024). Kidney structures in the AOAA and PAG groups were also better, compared to those in the IR control group (for AOAA, p=0.048; for PAG, p=0.024) (Fig. 4).
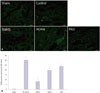
Apoptosis evaluation
Kidney sections from the IR control group showed increased TUNEL-positive cells, compared with those from the sham group (p=0.008). The degree of apoptosis in the NaHS group was significantly decreased, compared to that in the IR control group (p=0.024). Rats in the AOAA and PAG groups exhibited decreased TUNEL-positive cells, compared with those in the IR control group (for AOAA, p=0.024; for PAG, p=0.024) (Fig. 5).
DISCUSSION
This study demonstrates that exogenous H2S can attenuate renal IR injury. Its renoprotective effects were characterized not only by decreased serum BUN and Cr levels but also by attenuated histological damage that includes apoptosis caused by renal IR injury. Administration of NaHS in rats decreased MDA levels and preserved SOD levels, indicating reduced lipid peroxidation and preserved antioxidant enzymes, respectively. In addition, NaHS prevented renal IR-induced phosphorylation of MAPK family proteins (ERK, JNK, and p38). In the AOAA and PAG groups, renal function indices and MAPK phosphorylation levels did not change, compared to those in the renal IR control group, indicating that CGL or CBS inhibition had no effect on renal impairment of rats subjected to renal IR injury.
H2S has been identified as an important gaseous transmitter for molecular therapy in renal IR injury.1319 It was initially known as an environmentally toxic molecule; however, it is now recognized as a physiological molecule and third gasotransmitter, after NO and CO, in the mammalian body. Gasotransmitters exhibit various biological functions, including vasodilatation, angiogenesis, metabolic modulation, anti-oxidant activity, anti-inflammation, and anti-apoptotic activity, in various animal models.2021 Among many physiological and cytoprotective actions of H2S, one of the main mechanisms underlying H2S renoprotection is its anti-oxidative role. Several previous studies have demonstrated that H2S upregulates anti-oxidant defense, particularly under severe chronic conditions.2223 Moreover, H2S functions as a direct scavenger of ROS, thus preventing IR injury.2425 In this study, NaHS-treated rats exhibited significantly decreased serum BUN and Cr levels and improved renal tissue damage, compared to those in the IR control group. In addition, NaHS-treated rats exhibited decreased MDA levels, demonstrating its cytoprotective effect against oxidative stress. Increased SOD levels suggested that NaHS maintains the intracellular anti-oxidant capacity of kidney cells. Additionally, TUNEL-positive cells were decreased in the NaHS group, suggesting that NaHS offered a protective effect against renal tubular apoptosis. During IR injury, increased ROS initiates transition of mitochondrial permeability, which in turn induces pro-apoptotic molecules and stimulates the formation of various inflammatory cytokines.26 Therefore, decreased MDA and preserved SOD levels, along with reduced apoptosis, in renal cells might underlie the protective effect of NaHS on oxidative stress.
Three catalytic enzymes, CGL, CBS, and 3-MST, are associated with the production of endogenous H2S. CGL and CBS are primarily located on renal proximal tubules, and these are the main enzymes for H2S generation in the kidney.27 Kidney IR injury induces tubular epithelial cell damage, indicating that this might lead to impaired production of H2S. Han, et al.28 showed that the production of H2S was decreased in renal IR injury. Thus, H2S donor, NaHS, exerted the protective effects on the recovery from kidney damage.1328 CGL and CBS activities were reduced in various renal IR injury studies,1228 and inhibition of either enzyme by pharmacological inhibitors aggravated renal damage.1328 However, Roy, et al.29 showed that the inhibition of either of the enzymes (CGL or CBS) had no significant effect on the vascular system, whereas simultaneous inhibition of both the enzymes reduced H2S production. This is consistent with our results, which suggested that suppression of either CGL or CBS did not have any effect on renal IR. Therefore, we suspect that these two enzymes may have an interdependent regulation or that they may complement each other in maintaining the production of H2S. A recent study by Cortese-Krott, et al.30 demonstrated that the H2S and NO signaling pathways intertwine with coupled chemical reactions to generate three products, nitrosopersulfide, dinitrososulfide, and polysulfides, each with a specific chemical bioactivity. In this study, considering the response of both the inhibitors, we assume that some unknown interactions between H2S and other molecules, such as NO, an important mediator of IR injury, may affect the action of CGL or CBS inhibitors during renal IR injury. In addition, in the CGL or CBS group, there were fewer histological changes, compared to the IR injury control group, while there were no differences in the MDA levels. This discrepancy might be attributed to the nature of the reaction in histologic pathogenesis, such as inflammatory response, neutrophil activation, and angiogenesis. Meanwhile, anti-oxidant activity (SOD) was increased in the CGL or CBS group, which might have affect mild histological changes therein. Therefore, the exact role of these enzymes in renal IR injury is unclear, and further investigations will be needed in future.
Oxidative stress by IR injury can activate MAPK signaling. The MAPK signaling pathway play a central role in transferring extracellular signals to the nucleus.1 Among three MAPK subgroups, ERK is generally activated by growth factors and contributes to cell survival. Conversely, JNK and p38 are preferentially activated by environmental stress and contribute to cell death.3132 However, these roles are not always distinctive, and cell fate associated with MAPK pathways may be determined by the nature of the environmental stress, activation duration, cell type, or the activities of other signaling pathways.3334 The influence of MAPK on the cytoprotective effect of H2S has been extensively studied. The neuroprotective effect of H2S has been reported to be involved in the suppression of ERK activation.35 Further, activation of ERK, JNK, and p38 has been shown to be associated with chemical hypoxia-induced neuronal injury and with H2S-protected neuronal injuries via ROS-activated ERK and p38 MAPK signaling pathways.36 In addition, NaHS treatment has been found to attenuate the phosphorylation of ERK, JNK, and p38 in renal IR injury.18 Here, we confirm that renal ischemia for 30 min and reperfusion for 24 h results in activation of ERK, JNK, and p38. Above all, we have shown that these effects are reduced by exogenous H2S treatment.
The present study has certain limitations. Here, we focused only on oxidative stress in renal IR injury. In addition to oxidative stress, several other events, such as inflammatory responses and neutrophil activation, have also been implicated in the pathognomonic process. Furthermore, we tried to identify the relative contribution of the enzymes involved in H2S production by individual inhibition of AOAA or PAG; however, combined inhibition of both should also be investigated to reach a definite conclusion on H2S production.
In summary, we have demonstrated that exogenous H2S mitigates renal damage caused by IR injury. In addition, its proposed beneficial effect is, in part, due to anti-oxidative stress mediated via modulation of MAPK signaling. Therefore, the present study suggests that exogenous H2S may be beneficial in a variety of conditions associated with renal IR injury.



 XML Download
XML Download