

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Atopic march (AM) is the progression from atopic dermatitis (AD) to allergic rhinitis and asthma. The development of AD is as high as 20% of children worldwide and continues to increase1. AD seems to be caused by both genetic and environmental factors, and is correlated with a family history of allergic disease2. AD, asthma, and allergic rhinitis share an immune response that includes T helper type 2 (Th2) cell-mediated cellular components and serum immunoglobulin (Ig)E levels associated with allergic inflammation3. Whole exome sequencing (WES) has recently been used to successfully identify causal variants by filtering big data in cancer and Mendelian and rare disorders4. Common and complex diseases often have strong heritable components, and the identification of genetic variants in complex disorders is crucial to understanding their evolutionary background56.
Thymic stromal lymphopoietin (TSLP) is an important prime protein which is involved in the development of allergic asthma and rhinitis. The expression of TSLP mRNA is higher in human bronchial epithelial cells and primary skin keratinocytes compared with control subjects. The differentiation of naïve T cells into Th2 cells is promoted by TSLP, which also supports the maturation of antigen-presenting cells. These factors are thought to be pathogenic in allergic diseases. Overexpression of transgenic murine TSLP in keratinocytes developed a spontaneous AD-like phenotype7. The removal of TSLP signaling reduced the expression of cytokines (interleukin [IL]-4, IL-13) produced by Th2 lymphocytes, and serum IgE level in the allergic asthma responses8. Emerging data now suggest that epithelial cell-derived cytokines such as TSLP, IL-13, and IL-25 may drive the progression from atopic dermatitis to asthma and food allergy9. Consequentially, TSLP gene plays an important risk factor for the induction of allergic disease and progression of the allergic march81011. Polymorphisms of TSLP associated with allergic disorders have been reported in ethnic groups of various countries. A recent study demonstrated that genetic variations of TSLP can affect TSLP biological functions12. However, the distribution of TSLP polymorphisms in Koreans with AD or progressing in AM is not known. In this study, we observed polymorphisms of TSLP in Koreans with AD or AM using WES.
MATERIALS AND METHODS
Patients
This study included a control group of 20 subjects with only AD and AM patients for exome sequencing. AD patients who were adults with an onset age older than 18 years were recruited. Because these adult-onset patients did not have long-term follow-up since childhood, the presence of early-onset of AD relied on their family and patients' memory. The development of AD in adulthood significantly reduces the probability of the occurrence of asthma and allergic rhinitis compared to early-onset AD in infancy13. To validate candidate haplotype blocks between AM and AD in Koreans, we performed Sanger sequencing in 60 AD and 59 AM patients. Clinical information was displayed in Supplementary Table 1. This study was reviewed and approved by the Chung-Ang University Hospital Institutional Review Board, IRB no. C2015258 (1716). Informed consent was submitted by all subjects when they were enrolled. All groups were diagnosed with AD and AM by a dermatologist.
Whole-exome sequencing
Isolation of human genomic DNA from the peripheral blood was performed using a QIAamp DNA Mini Kit (Qiagen Inc, Valencia, CA, USA). The DNA quality and quantity were assessed with a Nanodrop spectrometer (Nanodrop Technologies, Wilmington, DE, USA) and a Qubit fluorometer (Life Technologies, Grand Island, NY, USA). WES was carried out using SureSelect Human All Exon V4+UTR 71 Mb (Agilent, Santa Clara, CA, USA), following the manufacturer's standard protocol. Genomic DNA was sheared using the g-TUBE™ (Covaris, Woburn, MA, USA). A paired-end DNA sequencing library was prepared through shearing, end-repair, A-tailing, peak detection, PE adaptor ligation, and amplification. After the library was hybridized with bait sequences for 24 hours, it was purified and amplified with an index barcode tag, and the library quality and quantity were determined. The exome library was sequenced with the 100-bp paired-end mode of the HiSeq SBS kit.
Sanger sequencing
Polymerase chain reaction (PCR) amplification of two single nucleotide polymorphisms (SNPs) was performed at 95℃ for 10 minutes, followed by 35 cycles at 95℃ for 30 seconds, 55℃~58℃ for 30 seconds and 72℃ for 40 seconds, with a final extension at 72℃ for 1 minute 30 seconds. The PCR reaction mixtures (total volume 50 µl) contained 25 µl of 2X EF-Taq premix (SolGent, Seoul, Korea), 18 µl of distilled water, 2.5 µl of oligonucleotide primer (10 pmol/µl), and 2 µl of template containing 20 ng genomic DNA. The PCR products were purified using a PCR purification kit (Favorgen, Pingtung, Taiwan) and were sequenced on an Applied Biosystems 3500 DNA sequencer (Foster City, CA, USA) according to the manufacturer's instructions.
Statistical analysis
Using logistic regression analysis, crude odds ratios (ORs), and 95% confidence intervals (CIs), associations between AM and AD with the SNPs under investigation were calculated. Pairwise Linkage disequilibrium was calculated as D′/LOD and r2, and Hardy-Weinberg Equilibrium and case-control association test were examined using Haploview software version 4.2 (http://www.broad.mit.edu/mpg/haploview/)14.
RESULTS
This study included a control group of 20 subjects with only AD (27.1±8.6 years old) and AM patients (24.9±6.0 years old) for exome sequencing. The clinical phenotype of the 20 recruited AM patients included 19 with AD and allergic rhinitis and one with AD and asthma. The AD control and AM groups did not differ significantly in age or blood eosinophil count. Serum total IgE levels were two times higher in the AM group than in AD controls (Supplementary Table 2).
We detected a total of nine SNPs of TSLP in the Korean subjects. These SNPs did not deviate from Hardy-Weinberg equilibrium (p>0.05; Table 1). Four (rs139817258, rs11466749, rs11466750 and rs10073816) of the nine SNPs were found in exon4 (3′ UTR): c.*556G>A, c.*813A>G, c.*1122G>A and c.*1717G>A. Another two SNPs (rs2289277 and rs2289278) were in intron2: c.217-142C>G and c.217-61C>G. The other two SNPs (rs3806933 and rs2289276) were in the promoter: c.-847C>T and c.-82C>T. The last one (rs191607411) was an upstream variant: c.-1453C>T. rs191607411 and rs139817258 were identified as rare variants, with global minor allele frequencies (MAFs) of 0.0012 and 0.0040, respectively, although the Korean MAFs were higher. The other seven variants were common with Global and Korean MAFs greater than 5%. Patients with genotypes of a heterozygote and homozygote minor allele were counted in the AD and AM groups (Table 1).
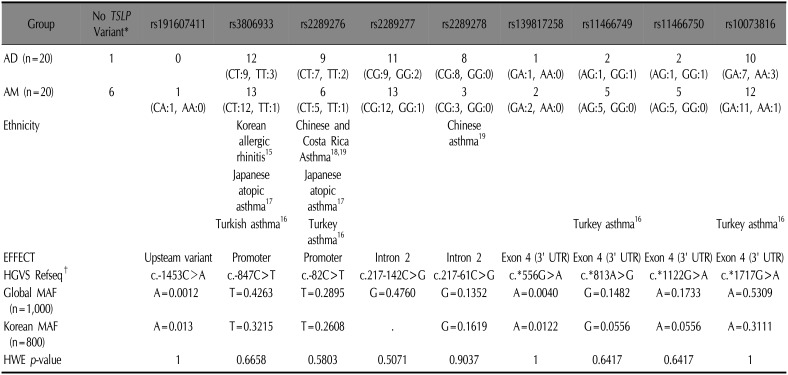
Table 1
Loci and frequency of TSLP variants detected in AD and AM using WES
| Group | No TSLP Variant* | rs191607411 | rs3806933 | rs2289276 | rs2289277 | rs2289278 | rs139817258 | rs11466749 | rs11466750 | rs10073816 |
|---|---|---|---|---|---|---|---|---|---|---|
| AD (n=20) | 1 | 0 | 12 (CT:9, TT:3) | 9(CT:7, TT:2) | 11 (CG:9, GG:2) | 8 (CG:8, GG:0) | 1 (GA:1, AA:0) | 2 (AG:1, GG:1) | 2 (AG:1, GG:1) | 10 (GA:7, AA:3) |
| AM (n=20) | 6 | 1 (CA:1, AA:0) | 13 (CT:12, TT:1) | 6 (CT:5, TT:1) | 13 (CG:12, GG:1) | 3 (CG:3, GG:0) | 2 (GA:2, AA:0) | 5 (AG:5, GG:0) | 5 (AG:5, GG:0) | 12 (GA:11, AA:1) |
| Ethnicity | Korean allergic rhinitis15 | Chinese and Costa Rica Asthma1819 | Chinese asthma19 | |||||||
| Japanese atopic asthma17 | Japanese atopic asthma17 | |||||||||
| Turkish asthma16 | Turkey asthma16 | Turkey asthma16 | Turkey asthma16 | |||||||
| EFFECT | Upsteam variant | Promoter | Intron 2 | Intron 2 | Exon 4 (3' UTR) | Exon 4 (3' UTR) | Exon 4 (3' UTR) | Exon 4 (3' UTR) | ||
| HGVS Refseq† | c.−1453C>A | c.−847C>T | c.−82C>T | c.217−142C>G | c.217−61C>G | c.*556G>A | c.*813A>G | c.*1122G>A | c.*1717G>A | |
| Global MAF (n=1,000) | A=0.0012 | T=0.4263 | T=0.2895 | G=0.4760 | G=0.1352 | A=0.0040 | G=0.1482 | A=0.1733 | A=0.5309 | |
| Korean MAF (n=800) | A=0.013 | T=0.3215 | T=0.2608 | . | G=0.1619 | A=0.0122 | G=0.0556 | A=0.0556 | A=0.3111 | |
| HWE p-value | 1 | 0.6658 | 0.5803 | 0.5071 | 0.9037 | 1 | 0.6417 | 0.6417 | 1 |
![]()
Seven of our patients had none of the nine TSLP SNPs detected by WES: The seven comprised six of the 20 in the AM group and one of the 20 in the AD group. The allele and genotype frequencies of the nine SNPs were assessed between the AD (controls) and AM (cases) groups (Supplementary Table 3). Variant risk factors for AM were estimated with ORs and 95% CIs. Individually, none of the nine SNPs was significantly associated with AD or AM. However, the absence of all nine SNPs was associated with a trend towards an enhanced risk of AM compared to AD with an OR of 8.14 (95% CI 0.8784~75.4825, p=0.0649) (Table 2).
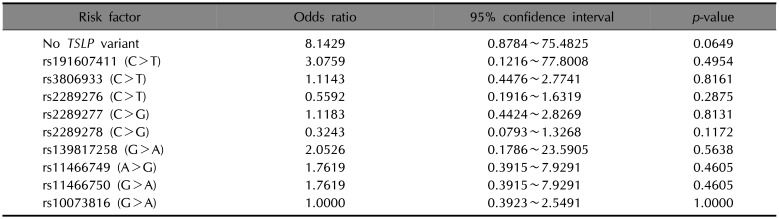
Table 2
Prevalence of risk factor and the allele frequencies in Atopic march (case) and atopic dermatitis (control), odds ratios and 95% confidence intervals
![]()
Three SNP pairs (rs3806933, rs2289276 and rs2289277) were in high linkage disequilibrium (LD, D′=1), as were another three SNP pairs (rs11466749, rs11466750 and rs10073816). Two haplotype blocks were created based on these triads. A strong LD score (D′=0.94) was identified between the two haplotype blocks (Supplementary Fig. 1A). The r2 value is presented in (Supplementary Fig. 1B). To assess the influence of the two blocks between AM and AD, a case-control association test was conducted with a chi-square test. There was a trend towards a difference with the TCG haplotype of block 1 (rs3806933, rs2289276 and rs2289277), (p=0.0769; Supplementary Table 4).
We next performed the validation of haplotype block 1 (rs3806933, rs2289276 and rs2289277) and haplotype block 2 (rs11466749, rs11466750 and rs10073816) in 60 AD and 59 AM Korean populations (Supplementary Table 1). The blocks were separated, and two SNP pairs (rs3806933 and rs2289276) and (rs11466749 and rs11466750) were in high linkage disequilibrium (LD), respectively (D′=0.97, D′=1) (Fig. 1). These haplotype blocks were assessed the frequency of each haplotype in AD and AM patients (Table 3). There was no association of the block 1 (rs3806933 and rs2289276) and the block 2 (rs11466749 and rs11466750) haplotypes between the AD and AM groups.
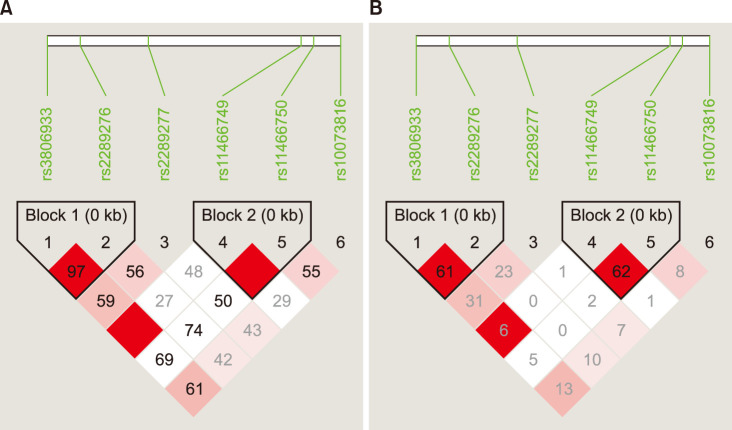
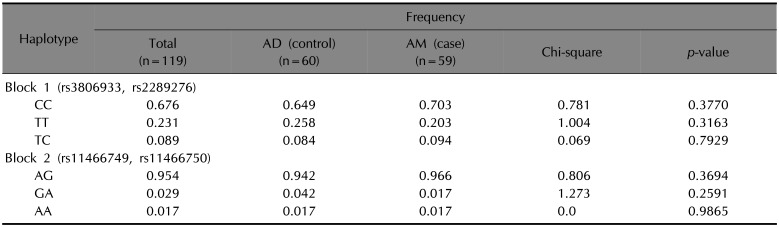
Fig. 1
Pairwise linkage disequilibrium (LD) map for six single nucleotide polymorphisms of thymic stromal lymphopoietin gene. (A) Pairwise D′/LOD and (B) r2 value are indicated as colors. White to red colors (closer to 1) representing the increasing strength of LD.
![]()
Table 3
Haplotype association analysis of TSLP polymorphisms in AD (control) and AM (case)
![]()
DISCUSSION
Some of nine detected SNPs have been correlated with susceptibility to allergic diseases in other studies. rs3806933, which is located in the promoter region, is a functional variant that enhances transcription factor activating protein (AP)-1 binding by creating a binding site12. It is significantly associated with susceptibility to Korean allergic rhinitis and Japanese and Turkish asthma151617. The high risk of Japanese asthma with an OR of 1.25 (95% CI 1.07~1.47, p=0.0063) was confirmed by the increase of a minor allele (T) frequency at rs3806933 in asthma patients compared to control group, whereas the low risk of Turkish asthma with an OR of 0.53 (95% CI 0.32~0.89, p=0.016) was observed with the increase of a major allele (C) frequency in asthma group1617. The frequency of a major allele (CC) genotype at rs3806933, the AA genotype at rs11466749 and GG genotype at rs10073816 were significantly higher in a Turkish asthma group. The frequency of a major allele (CC) genotype at rs3806933 was also increased in Korean allergic rhinitis with an OR of 0.66 (95% CI 0.50~0.88, p=0.015)15. The opposite results were observed in different ethnic group. AM patients of our data included 19 with AD and allergic rhinitis and one with AD and asthma.
The global MAF indicates that rs2289276 is a common variant located in the promoter region. It is also related to Costa Rican, Japanese, and Turkish asthma161718. Rs2289276 potentially increases its transcriptional activity by altering the binding affinity of AP-2α, which is a transcription suppression factor17. A minor allele (T) frequency as a risk factor was elevated at rs2289276 in Chinese and Japanese asthma groups1719. In contrast, a major allele (C) frequency was significantly elevated in Turkish asthma. The CC genotype of rs2289276 is significantly associated with higher eosinophil numbers and lower forced expiratory volume (FEV1) in Turkish asthmatics16. In a case control study involving a Chinese Han population, the major C allele of rs2289278 was associated with reduced FEV1:forced vital capacity (FVC)19.
Significant differences in rs11466749 and rs10073816 frequencies between healthy children and asthmatics were observed in a Turkish study16. rs191607411, rs2289277, rs139817258, and rs11466750 have not been previously identified in studies related to allergy.
FLG mutation disrupts skin barrier function, leading to more sensitization and greatly increasing the risk of AD20. TSLP is highly expressed in the epidermal keratinocytes and plays a key role in immunological events in atopic march. Thus, the impairment of inflammatory cytokines through genetic mutation may help to prevent allergic disease. The rs2289276 and rs3806933 located in the promoter region enhance transcriptional activity17. However, it is difficult to conclude the risk of TSLP variants because of the opposite results in different ethnic group.
We found that Korean patients without the nine TSLP variants had an 8.14 times higher risk of progressing from AD to AM. Although the OR did not reach statistical significance, there was a trend toward significant (p=0.0649). Thus, the increase of major allele frequency in each TSLP variant increase the risk of AM. We need to investigate the newly discovered rs191607411, rs2289277, rs139817258 and rs11466750 variants in Koreans. These variants are located in the 3-prime untranslated region (3′ UTR) of exon 4. The 3′ UTR contains regulatory regions including polyadenylation, translation, localization, and mRNA stability. They have critical functions which can influence the fate of mRNA21. Thus, there may be effects between mutations. Although our study was limited by a small sample size, the grouping of nine TSLP polymorphisms detected in AD and AM Koreans tells a novel story. Two haplotype blocks were identified with high LD (D′=1). To overcome small sample size and confirm a coinherited tendency of haplotype block, additional case-control study was conducted about two candidate SNP block1 (rs3806933, rs2289276 and rs2289277) and SNP block2 (rs11466749, rs11466750 and rs10073816) in 60 AD and 59 AM Korean populations. Unexpectedly, each haplotype block 1 (rs3806933, rs2289276 and rs2289277) and haplotype block 2 (rs11466749, rs11466750 and rs10073816) of three SNP pairs became separated in two piece. However, the significant coinherited feature (D′=0.97, D′=1) was identified in each two SNP pairs (rs3806933 and rs2289276) and (rs11466749 and rs11466750). Rs3806933 and rs2289276 variants were already verified the biological functionality in vitro, respectively17. There was no association between AD and AM for these two SNP blocks (rs3806933 and rs2289276) and (rs11466749 and rs11466750). Thus, the genetic clues contributed to processing allergic march was not detected in haplotype association test. We need to compare control groups in future study.
The finding of nine TSLP polymorphisms and haplotypes in Koreans with AD or AM is novel. Five of the nine SNPs detected in Korean patients have also been associated with Chinese, Japanese, Turkish, Costa Ricans. Our findings indicate the increased risk of AM in patients without the nine TSLP compared to AD. The data demonstrate one functional haplotype blocks together with the other candidate block are observed in Koreans.
We suggest that the increase of major allele frequency of respective TSLP variants may enhance the risk of AM. These data will contribute to an improved genetic surveillance system in the early diagnosis technology of allergic disease.
 XML Download
XML Download