

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Diabetes mellitus (DM) is a metabolic disorder characterized by defective insulin secretion and hyperglycemia. Numerous patients with diabetes develop associated complications, such as cardiovascular disease, diabetic neuropathy, and kidney disease [1234]. In addition, one of the main complications of diabetes affecting quality of life is impaired bladder function, which is experienced by 50% of diabetic patients [5]. The symptoms of diabetic bladder dysfunction include increased residual urine volume after voiding and impaired detrusor contractility [67]. Approximately 23% of diabetic patients have reduced bladder contractility [8].
The urinary bladder is an elastic organ that stores urine [9]. In the bladder, the smooth muscle of the detrusor can both contract and relax to control the excretion of urine [10]. Muscarinic receptors are principally responsible for mediating bladder contraction with M2 muscarinic receptors making up about 90% of the total muscarinic receptors in bladder smooth muscle in rats [1112]. Electrical field stimulation (EFS) can trigger the release of cholinergic neuronal mediators and induce contractions through muscarinic receptors in bladder smooth muscle [131415]. Specifically, EFS-induced contraction is mediated mainly by acetylcholine (Ach) release from parasympathetic nerves. There are several types of receptors present in rat bladders, including adrenergic and purinergic receptors [161718]. Nitric oxide synthase and phosphodiesterase type 5 (PDE5) are also involved in bladder contractility [1920]. Hydrogen sulfide (H2S) can also induce contraction of bladder smooth muscle and H2S synthetic enzymes are produced in the bladder [212223]. It has been reported that PDE5 inhibitors increase contractile force in normal rat bladders [20]. Nitric oxide (NO) activates guanylate cyclase, which results in increased levels of cyclic guanosine monophosphate (cGMP), thus leads to relaxation of smooth muscle. PDE5 inhibitors stimulate relaxation of bladder smooth muscle by inhibiting cGMP degradation by PDE5.
A decrease in bladder contractility has been reported in rats with diabetes mellitus (DM rats) [24], in which a hypocontractile state has been reported, especially in animals aged 12–20 weeks [25]. This decrease in bladder contractility occurs for a variety of reasons. One is a decrease in sensitivity to ACh in DM rats [2627]. Another is changes in calcium sensitivity that have been suggested to be related to changes in bladder contraction [2829]. In addition, NO production increases in hyperglycemia and diabetic cystopathy [3031]. Taken together, these data suggest that G protein-coupled receptors (GPCRs), intracellular signaling pathways, and NO production play roles in bladder contractility in DM rats.
Using specific pharmacological inhibitors, the mechanisms underlying changes in bladder smooth muscle contraction in DM rats were identified. The pharmacological inhibitors used in this study were the muscarinic receptor antagonist atropine, the phospholipase C (PLC) inhibitor U73122, the adenosine A1 receptor antagonist DPCPX, the PDE5 inhibitor udenafil, the α1-receptor antagonist prazosin, the smooth muscle relaxer NaHS, the calcium channel blocker verapamil, and the protein kinase C (PKC) inhibitor chelerythrine. Contractions were induced electrically using electrical field stimulation (EFS) and were compared between normal rats and rats with streptozotocin (STZ)-induced DM. Therefore, the aim of the present study was to investigate the role of GPCRs, intracellular signaling pathways, and NO production in bladder smooth muscle contractility in DM rats.
METHODS
Animals
Male Sprague-Dawley (SD) rats weighing 250–280 g were supplied by Samtako Bio (Osan, South Korea). The animals were housed in groups in cages with wire-net floors in a temperatureand humidity-controlled room (24–25℃ and 70–75%, respectively) with a 12-h light-dark cycle. Rats were fed a normal laboratory diet supplied by Samtako Bio (Osan, South Korea) and fasted for 24 h prior to the experiment. The experiments were performed in accordance with the guidelines and approval of the Institutional Animal Care Use Committee of Chung-Ang University (IACUC 2017-00072).
Drugs and chemicals
Udenafil, atropine, prazosin, NaHS, verapamil, U73122, DPCPX, chelerythrine, and STZ were all purchased from Sigma (St. Louis, MO, USA). The Krebs buffer consisted of 133 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2, 1.35 mM NaH2PO4, 0.6 mM MgSO4, 16.3 mM NaHCO3, and 7.8 mM dextrose. Unless otherwise noted, it was purchased from Sigma (St. Louis, MO, USA).
Induction of diabetes
STZ is a chemical that is particularly toxic to insulin-producing beta cells in the pancreas and is often used in medical research in animal models of hyperglycemia and diabetes. Experimental diabetes was induced in overnight-fasted rats using a single dose (65 mg/kg) of STZ (Sigma). STZ was dissolved in normal saline and then administered intraperitoneally to the rats. The blood glucose levels of rats were checked two days after STZ administration using a CareSens II glucose meter from i-SENS (Seoul, South Korea). Rats with blood glucose levels higher than 300 mg/dl were considered to have DM.
Organ bath experiment using bladder strips
Two weeks after STZ administration, the rats were sacrificed using CO2. The bladders were removed and placed in Krebs solution. The bladders were then sliced longitudinally into equal strips (2 mm×7 mm). A mixture of oxygen and CO2 gas was supplied continuously to the strips. Transversely-oriented muscle strips were taken from the rat bladders. The strips were then cut into 2–3 minor strips and silk ligatures were tied to both ends. The muscle strips were then mounted in separate 1 ml muscle chambers with one wire fixed to the bottom of the muscle chamber and the other was attached to a force transducer (FT03 Grass Instruments Co., Quincy, MA, USA). Changes in isometric force were recorded on a polygraph (Grass model 79, Quincy, MA, USA). The muscle strips were initially stretched using 1 g to generate conditions for optimal force development and were equilibrated for over 60 min while continuously being perfused with oxygenated Krebs solution.
EFS
The bladder strips were stimulated with 10 s pulse trains 40 V in amplitude with pulse durations of 1 ms at frequencies of 2-10 Hz using a stimulator (Model S 88, Grass Instruments) through platinum wire electrodes placed longitudinally on either side of the strips. After a stable resting tone of muscle strips was obtained, the frequency-response relationship (2–10 Hz) was studied, where the strips were washed five times and allowed to equilibrate for 30 mins after EFS to permit the strips to completely recover.
Data analysis
The contraction response to EFS is expressed as percentage (%) versus control. Data are expressed as mean±standard error of the mean. Statistically significant differences among the groups were identified using two-way repeated measures ANOVA. A p-value of less than 0.05 indicated statistical significance.
RESULTS
Comparison of bladder smooth muscle tension between normal and DM rats
Representative tracings of tension in normal and DM rats are presented in Figs. 1A and B. Contractions were induced using EFS (2–10 Hz). The average bladder contractilities upon EFS were compared between the normal and DM rat curves and analyzed using two-way ANOVA. In DM rat muscle, there was a significant decrease in EFS-induced contractility compared to that in normal rat muscle.
GPCR-related signaling on EFS-induced bladder smooth muscle contraction
As shown in Fig. 2, pretreatment with the muscarinic receptor antagonist atropine (10−6 M) inhibited EFS-induced contraction of bladder smooth muscle from both normal and DM rats. The percentages of contraction were 62% in the normal group and 58% in the DM group compared to that of the controls.
The effect of α1-adrenoceptor antagonist prazosin (10−6 M) on EFS-induced contraction of bladder smooth muscle is shown in Fig. 3. Rat bladder smooth muscle was pretreated with prazosin and contractility was measured. After pretreatment, there was a significant difference between contraction of muscles from normal and DM rats in response to EFS, where the percent contraction compared to the control was higher for DM rat muscle than for normal rat muscle.
As shown in Fig. 4, there was no difference between the normal and DM rat muscles when the muscle strips were treated with the adenosine A1 receptor antagonist DPCPX (10−6 M). In addition, there was no difference between the controls. Experiments examining the roles of GPCR-mediated signaling pathways revealed a significantly different role for a1 adrenoceptors in contraction responses between normal and DM rat muscles.
Effect of inhibition of intracellular signaling pathways on EFS-induced contraction of bladder smooth muscle
The effect of the calcium channel blocker verapamil (10−6 M) on bladder smooth muscle contraction is shown in Fig. 5A. Following treatment with verapamil, contractions in response to EFS decreased in both normal and DM rat muscles. The magnitude of this decrease in the contractions was larger in normal rat muscles than in DM rat muscles.
The effect of the PLC inhibitor U73122 (10−6 M) on bladder smooth muscle contraction is shown in Fig. 5B. Following treatment with U73122, there was a significant difference between DM and normal rat EFS-induced muscle contraction, where the percent contraction in DM rat muscles was lower than that in normal rat muscles.
The effect of the PDE5 inhibitor udenafil (10−6 M) on bladder smooth muscle contraction is shown in Fig. 6A. A significant difference was observed between normal and DM rat muscles following udenafil treatment, where the DM rat muscles had a higher percent contraction than normal rat muscles.
As shown in Fig. 6B, there was no difference in EFS-induced contractions between normal and DM rat muscles when the smooth muscle relaxer NaHS (10−6 M) was used to treat the muscle strips. In addition, there were no significant differences between the controls.
As shown in Fig. 7, there were no differences in EFS-induced contraction between normal and DM rat muscles following treatment with the PKC inhibitor chelerythrine (10−6 M). In addition, there were no differences between the controls.
In these experiments examining intracellular signaling pathways, modulators of calcium channels, PLC, and PDE5 were found to have significantly different effects on normal and DM rat bladder smooth muscle contraction in response to EFS.
A summary of the changes in EFS-induced contraction of DM rat bladder muscle following treatment with different agents is presented in Fig. 8. GPCR- and NO-mediated intracellular signaling pathways are involved in changes in the contraction or relaxation of bladder smooth muscle. In DM rats, an alteration in GPCR-mediated PLC signaling affected IP3-dependent intracellular Ca2+ release and PDE5 activity affected contraction or relaxation of bladder muscle in DM rats.
DISCUSSION
Bladder dysfunction is one of the main complications occurring in DM [32]. A decrease in detrusor contractility is a common symptom of bladder dysfunction [7]. Many different factors influence bladder smooth muscle contraction. The present study was conducted to identify receptors and/or enzymes with roles in contraction of bladder smooth muscle in a rat model of DM [33].
In this study, atropine decreased contractility in normal and DM rat bladder muscles to a similar extent. By contrast, treatment with prazosin (an α1-receptor antagonist), udenafil (a PDE5 inhibitor), verapamil (a calcium channel blocker), and U73122 (a PLC inhibitor) yielded significant differences in contractions in response to EFS between normal and DM rat muscles.
Role of GPCRs in bladder contraction in DM
To investigate the role of GPCRs in bladder contraction in DM, atropine, prazosin, and DPCPX were used as pharmacological tools. The α1-adrenergic receptors, especially the α1A-adrenoceptor subtype, are expressed in rat bladders. Agonists acting on α1-adrenoceptors enhance EFS-induced release of ACh in isolated rat bladders. It has also been reported that α1-adrenoceptor agonists produce contractions that are about 10–43% of those achieved using muscarinic stimulation [34]. In addition, an enhanced adrenoceptor receptor response has been observed in DM rat bladders [35]. These data suggest that α1-adrenoceptors may have a role in bladder contraction in DM.
Adenosine A1 receptors are expressed in the uroepithelium of the bladder [36]. DPCPX (8-cyclopentyl-1, 3-dipropylxanthine) acts as a selective antagonist of the adenosine A1 receptor [3738]. These adenosine A1 receptors can activate PLC, increasing cytoplasmic Ca2+ concentrations, as well as inhibit adenylyl cyclase [3940]. ATP can also induce contractions through activation of P2X purinergic receptors [41]. Adenosine-induced relaxation is thought to primarily occur through P1A1 receptors because treatment with an antagonist of the P1A1 receptor (DPCPX) decreases adenosine-induced relaxation in normal rat bladder [18]. In this study, prazosin had a significantly different effect on contraction in normal and DM rat muscles.
Changes in intracellular signaling pathways in smooth muscle bladder contraction in DM rats
Udenafil is a PDE5 inhibitor used for the treatment of erectile dysfunction. Interestingly, mRNA encoding PDE5 has been found in rat bladders [42]. It has also been reported that treatment with vardenafil, another PDE5 inhibitor, increases the contractility of bladder smooth muscle, potentially through changes in cGMP that lead to increases in intracellular calcium levels and thus increase bladder contraction [43]. The present study also suggests that the cGMP pathway may be involved in the control of relaxation of bladder smooth muscle in DM.
The density of muscarinic receptors in rat bladders increases during the early stages of DM [44] and M3 muscarinic receptors cause bladder contraction through a PLC-independent mechanism [45]. The main pathway for muscarinic stimulation of contraction of bladder smooth muscle involves activation of PLC, which leads to the generation of inositol-1, 4, 5-trisphosphate (IP3) [10]. IP3 induces Ca2+ release and leads to bladder contraction [46]. Numerous studies have observed alterations in calcium channel sensitivity and calcium levels in DM rats. In particular, it has been shown that intracellular calcium levels are elevated in animal models of DM [282947].
NaHS induces relaxation of bladder smooth muscle and H2S synthetic enzymes have been shown to be expressed in the bladder. However, we found no significant difference between normal and DM rat bladder smooth muscle contraction following NaHS treatment.
Several studies have investigated the effect of DM on contraction of cardiac muscle and colonic smooth muscle. It has been reported that the cholinergic response and contractility of colonic smooth muscle are reduced in DM rats. The contractile response of the proximal colon following treatment with carbachol, an ACh receptor agonist, was also found to be significantly weaker in DM rats. [48]. Another study on DM using female GK rats found reduced contraction of left ventricular muscle [49].
Our study has several limitations. First, only male rats were used. Male rats have organs not present in female rats, such as the penis and prostate. These male-specific organs may have effects on bladder smooth muscle contraction that we did not take into account. Second, the effect of age was not studied as all the rats used in this study were of a similar age (10 weeks). In this regard, it has been reported that the contractility of rat muscle changes with age [25]. Third, a hypercontractile state was also observed in DM rats. There might be various other factors affecting contraction of bladder smooth muscle. Accordingly, additional experiments will be required to address these limitations.
In conclusion, the present study found that bladder contractility was decreased in DM rats. Although atropine did not inhibit contraction of DM rat muscles, the effects of a PLC inhibitor, a calcium channel blocker, and a PDE5 inhibitor were significantly different between normal and DM rat muscles. Overall, decreased contractility of DM rat muscles was a result of alterations in PDE5 and PLC activity, including changes in the PLC signaling pathway that mediates IP3-induced intracellular Ca2+ release.







 ) indicate signaling pathway differences between normal and DM rats. Phospholipase C generates the intracellular second messenger IP3 from the membrane lipid PIP2. IP3 then induces Ca2+ release and the released Ca2+ binds to calmodulin. Ca2+/calmodulin activates MLCK, which phosphorylates myosin light chain, thus leading to contraction. In bladder smooth muscles from DM rats, changes in G protein-coupled receptor-mediated phospholipase C signaling affects IP3-dependent Ca2+ release and changes in PDE5 affect contraction or relaxation. These factors are expected to result in a decrease in bladder contractility. DM, diabetes mellitus; EFS, electrical field stimulation; PIP2, phosphatidylinositol 4,5-bisphosphate; PLC, phospholipase C; DAG, diacylglycerol; IP3, inositol trisphosphate; CaM, calmodulin; MLCK, myosin light chain kinase; MLC-P, myosin light chain phosphatase; NO, nitric oxide; PKG, protein kinase G; PDE5, phosphodiesterase 5.
) indicate signaling pathway differences between normal and DM rats. Phospholipase C generates the intracellular second messenger IP3 from the membrane lipid PIP2. IP3 then induces Ca2+ release and the released Ca2+ binds to calmodulin. Ca2+/calmodulin activates MLCK, which phosphorylates myosin light chain, thus leading to contraction. In bladder smooth muscles from DM rats, changes in G protein-coupled receptor-mediated phospholipase C signaling affects IP3-dependent Ca2+ release and changes in PDE5 affect contraction or relaxation. These factors are expected to result in a decrease in bladder contractility. DM, diabetes mellitus; EFS, electrical field stimulation; PIP2, phosphatidylinositol 4,5-bisphosphate; PLC, phospholipase C; DAG, diacylglycerol; IP3, inositol trisphosphate; CaM, calmodulin; MLCK, myosin light chain kinase; MLC-P, myosin light chain phosphatase; NO, nitric oxide; PKG, protein kinase G; PDE5, phosphodiesterase 5.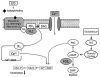
 XML Download
XML Download