

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Unlike other 5-hydroxytryptamine (5-HT) receptors, which are G protein-coupled receptors, 5-HT3 receptor is a ligand-gated ion channel and belongs to a Cys-loop receptor family which includes nicotinic acetylcholine receptors, γ-aminobutyric acid (GABA) type A receptors and glycine receptors [12]. The 5-HT3 receptor is permeable to cations, so it can induce a fast inward current in the neurons [3]. It mainly locates in presynaptic terminals, thus it could modulate the release of neurotransmitters such as dopamine, GABA, acetylcholine, substance P, and also 5-HT itself [45]. 5-HT3 receptors are widely distributed in the brain, relatively high in the area postrema and nucleus tractus solitarius. Also, it is found in the hippocampus, frontal cortex, cingulate cortex, amygdala, nucleus accumbens, substantia nigra and ventral tegmental area. In outsides of the brain, dorsal horn and dorsal root ganglia of spinal cord, lung, stomach, colon, kidney, and inflammation cells express the 5-HT3 receptor [6].
Clinically, 5-HT3 receptor antagonists like ondansetron are widely used as antiemetics [7]. Also, 5-HT3 receptor antagonist could be effective for the treatment of morphine and ethanol addiction because it reduced dopamine release induced by ethanol and morphine in animal studies [89]. Other researches revealed that ethanol potentiated 5-HT3 receptor function [1011], which suggest that the 5-HT3 receptor could be related to ethanol addiction. Schizophrenia, depression, anxiety disorder, irritable bowel syndrome, anorexia, cognitive disorder, Alzheimer's disease, inflammatory pain, neuropathic pain, and migraine are also suggested to be potential therapeutic targets of 5-HT3 receptor modulators [61213]. However, the pathophysiologic and pharmacologic role of the 5-HT3 receptor for the chronic pain and analgesic action are still need to be elucidated. Amitriptyline, one of the tricyclic antidepressant, has an affinity for serotoninnorepinephrine reuptake transporters, muscarinic acetylcholine receptors, 5-HT1A and 5-HT2A receptors, a-adrenergic receptors, histamine receptors [14]. It is effective to treat migraine, fibromyalgia, neuropathic pain, and irritable bowel syndrome, but its pharmacologic mechanism is still unclear [151617]. The clinical uses of amitriptyline overlapped with the potential therapeutic targets of 5-HT3 receptor modulators. However, a direct effect of amitriptyline on 5-HT3 receptor was not tested yet, although the past study showed amitriptyline inhibited chimeric 5-HT3 receptor and reduced 5-HT3 receptor-mediated cGMP formation [1819].
Thus, we studied the effect of amitriptyline on the 5-HT3 receptor and its mechanism using a whole cell patch clamp recording with a fast drug application system and compared the recorded data with the simulated macroscopic current. Based on our study, we could suggest an expanding of pharmacologic action of amitriptyline on various neurologic and psychiatric diseases.
METHODS
Cell culture
NCB-20 neuroblastoma cells, which used for studying 5-HT3 receptor [112021], were provided by Dr. Lovinger (National Institute on Alcohol Abuse and Alcoholism, USA). Cells were incubated in the culture flask filled with culture medium which contained 89% Dulbecco modified Eagle's medium, 10% fetal bovine serum, and 1% hypoxanthine aminopterin thymidine supplement. Cells were grown in an incubator maintained by 5% CO2 at 37℃, harvested by 0.25% trypsin-EDTA, and seeded to a treated culture dish 24–48 h prior to recording. Culture medium was exchanged to extracellular solution 1–2 h before recording and cells were transferred on cover glasses and moved to the recording chamber.
Electrophysiology
Extracellular solution was made by (in mM) 150 NaCl, 2.5 KCl, 2.5 CaCl2, 0.1 MgCl2, 10 D-glucose and 10 N-(2-hydroxyethyl)piperazine-N′-2-ethansulfonic acid (HEPES) and pH was adjusted to 7.4 with NaOH and osmolality adjusted to 340 mOsm/kg with sucrose. Intracellular solution consisted of (in mM) 140 CsCl, 2 MgCl2, 5 ethylene glycol bis(2-aminoethylether)-N,N,N′,N′-tetraacetic acid (EGTA), 10 HEPES and pH adjusted to 7.2 with CsOH, osmolality adjusted to 310 mOsm/kg with sucrose.
Whole-cell patch clamp technique was performed at room temperature (24℃) by using a Multiclamp 700B amplifier (Molecular Devices, Sunnyvale, CA, USA) with a Carl Zeiss Axiovert 135M microscope (Jena, Germany). Pipettes were made by borosilicate glass capillaries (1B150-4, World Precision Instruments, Sarasota, FL, USA) which pulled with a horizontal micropipette puller (P-97, Sutter Instrument, Novato, CA, USA). Pipette tip resistance was formed around 2.5 MΩ when it was filled with an intracellular solution.
Signals were filtered at 2 kHz, digitized at 10 kHz, and saved using Digidata 1322 and pClamp 10.0 software (Molecular Devices). Pipette capacitance was compensated before making whole cell configuration and whole cell capacitance and series resistance were compensated before drug application. All compensations were done by Multiclamp 700B Commander software (Molecular Devices). Currents were recorded at the −50 mV holding potential except when measured the current-voltage relationship.
Drug preparation and application
After achieving whole-cell configuration at the −50 mV holding potential, cells were lifted and moved toward the one side of theta glass pipette (Clark Borosilicate Theta, Warner Instruments, 2 mm outer diameter, 1.4 mm inner diameter, 0.2 mm septum thickness) which was pulled to an outer diameter of ~300 mm and continuously perfused with extracellular solution. The solution flow was driven by gravity from the reservoirs. It was controlled by a perfusion valve control system (VC-8, Warner Instruments, Hamden, CT, USA). While the cell was placed on the side of the extracellular solution flow, the agonist or tested drug were perfused on the other side of theta glass tubing. We turned on the VC-8 valve control system 5 s prior to every drug application to wash the space of theta glass pipette. Theta glass pipette was mounted with a piezoelectric translator (P-601 PiezoMove Z Acturator, PhysikInstrumente, Karlsruhe, Germany), which moved theta pipette laterally in ms time resolution. Therefore the cell could rapidly move to the drug side from the extracellular solution side. After the cell was exposed to the agonist for a programmed time, the cell was returned to the original side of the theta pipette perfused with the extracellular solution.
Data analysis, statistics, and simulation
Peak amplitudes of current were measured by Clampfit software (Molecular Devices). All time constants and slopes were calculated by the built-in statistical tools in Clampfit software. Peak current data were normalized to the 10 µM 5-HT current for statistical analysis. Desensitized currents were fitted by first order exponential function by using Clampfit software. Normalized concentration-peak amplitude data were fitted to sigmoid curve calculated by the four-parameter logistic equation using Prism 6.0 (GraphPad Software, San Diego, CA, USA).
For calculating EC50, equation (1) was used.

Bottom of the equation (1) is minimal response, top is maximal response, X is the logarithm of 5-HT concentration, and EC50 is concentration of half between top and bottom.
For calculate IC50, equation (2) was used.

Bottom of the equation (2) is minimal response, top is maximal response, X is the logarithm of amitriptyline concentration, and IC50 is concentration of half between top and bottom.
The mean and S.E.M value were calculated by Prism 6.0. Statistical significance was determined by p value <0.05, calculated by the Paired student's t-test, ANOVA, and Tukey's multiple comparisons test using Prism 6.0.
We formulated a differential equation of the 5-HT3 receptor kinetic models derived from a Michaelis-Menten equation, and it was solved by the fourth order Runge-Kutta method using our recorded data. Thus we could obtain rate constants of the kinetic model and simulated current. All this process was done by the Berkeley Madonna® software developed by Robert Macey and George Oster of the University of California at Berkeley.
RESULTS
5-HT3 receptor currents
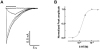
First, we studied the agonist concentration-response relationship using whole-cell patch clamp technique. Fig. 1A shows our representative 5-HT3 receptor currents of the NCB-20 cells depending on 5-HT concentrations of 0.3, 1, 3, 10, 30 µM. 5-HT was applied for 5 s, in 1 min interval at the −50 mV holding potential. As seen in Figs. 1A and B, the peak current was increased in a 5-HT concentration-dependent manner. A maximal current was obtained at 10 µM of 5-HT. Using equation (1), the calculated EC50 of peak currents by 5-HT was 1.73±0.08 µM with a Hill coefficient of 2.44±0.08 (n=10). The shape of current trace, EC50, and Hill coefficient values are all similar with our previous investigations [2223]. The currents induced by the application of 5-HT in NCB-20 cells were known to be mediated by the 5-HT3 receptor activation, because these currents were completely blocked by the GR 38032F and IC 205-930, the potent 5-HT3 receptor antagonists [24].
Effect of amitriptyline on 5-HT3 receptor currents
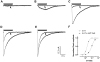
Figs. 2A and B present the 5-HT3 receptor current traces depending on amitriptyline concentration at the fixed 5-HT concentrations. To test the concentration-inhibition relationship, 3 µM or 10 µM 5-HT was applied concurrently with various concentration of amitriptyline (0.3, 1, 3, 10, 30 µM). Amitriptyline inhibited the 5-HT3 receptor currents in a concentration-dependent manner. The peak current was significantly blocked by 30 µM of amitriptyline (82.18±1.39% inhibition at 10 µM of 5-HT, p<0.001, n=10; 91.34±1.90% inhibition at 3 µM of 5-HT, p<0.001, n=10). In Fig. 2C, IC50 of amitriptyline on the 3 µM 5-HT-induced currents was 1.78±0.37 µM, and on the 10 µM 5-HT was 6.36±0.45 µM. The Hill coefficient of amitriptyline in 3 µM 5-HT was 1.00±0.08, and in 10 µM 5-HT was 0.91±0.04 (n=10 cells). IC50 and Hill coefficient were calculated by equation (2). Because the IC50 of the 3 µM 5-HT was smaller than 10 µM 5-HT (unpaired t-test, p<0.001, n=10), we expected that amitriptyline could block more effectively at a low concentration of 5-HT, which suggested that 5-HT concentration influence the inhibitory effect of amitriptyline.
Amitriptyline acts as a competitive blocker on 5-HT3 receptor
To elucidate how amitriptyline blocks the 5-HT3 receptor-mediated current, we tested whether amitriptyline acts as a non-competitive or competitive blocker on the 5-HT3 receptor. Because the IC50 of amitriptyline at 3 µM 5-HT was 1.78 µM, we used 3 µM of amitriptyline, above the IC50. We tested the effect of 3 µM amitriptyline co-applied with the 0.3, 1, 3, 10, 30 µM of 5-HT, and compared the peak current amplitudes with the 5-HT alone. Figs. 3A-E shows representative traces of the 5-HT3 receptor currents with or without amitriptyline. EC50 of 5-HT alone was 1.73±0.08 µM and Hill coefficient was 2.44±0.08 from the Fig. 1B. However, EC50 of the co-application of 5-HT and amitriptyline was 9.21±0.06 µM, and the Hill coefficient was 2.00±0.13 (Fig. 3F). Although EC50 was increased from 1.73 to 9.21 µM by amitriptyline (paired t-test, p<0.001, n=7), amitriptyline could not decrease the maximal peak current amplitudes induced by 30 µM of 5-HT (paired t-test, p=0.13, n=7). The normalized agonist concentration response curve was shifted to the right. Therefore, we could expect that amitriptyline acts as a competitive inhibitor on the 5-HT3 receptor.
Voltage independence of amitriptyline effect
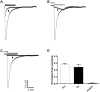
Voltage dependency of the 5-HT3 receptor current was performed to test the inhibitory mechanism of amitriptyline. We measured the 5-HT3 receptor-mediated current of 3 µM 5-HT with or without 3 µM amitriptyline at the holding potentials of −50, −30, −10, 0, 10, 30 mV, respectively. Figs. 4A and B show the current traces when we changed the holding potential from −50 mV to 30 mV. In Fig. 4C, the reversal potential in the absence of amitriptyline was 8.54±0.32 mV, and the reversal potential with amitriptyline was 8.88±0.44 mV. The difference between these reversal potentials were not statistically significant (p=0.4899, n=10).
Fig. 4D shows that the calculated inhibition ratios were not changed significantly over all the tested holding potential ranges. Because the result represents that amitriptyline works in a voltage independent manner, we could expect that amitriptyline does not act at the open pore of the receptor.
Comparison between pre-application and co-application of amitriptyline
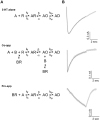
In order to test whether amitriptyline acts as a closed state blocker or not, we compared the effects of amitriptyline in different modes of application, because previous reports suggested that pre-application of a drug is effective when antagonist dominantly acts as a close state blocker [252627]. During co-application of amitriptyline (3 µM) with 3 µM 5-HT, the peak amplitude was decreased with shape of fast inward current which was similar with trace of 3 µM 5-HT alone (Fig. 5A). However, at the preapplication mode of amitriptyline, the 5-HT3 receptor-mediated current by 3 µM 5-HT raised with small rising slope to the end of 5-HT application without current decay (Fig. 5B). It was similar with the current of a lower concentration of 5-HT (i.e., ≤ 1µM, see Fig. 1A). However, we could not exclude the possibility that the slow activation observed in the pre-application experiments likely caused by washout of amitriptyline, which gradually increase the number of available receptors. When amitriptyline was pretreated and followed by co-application, the 5-HT3 receptor current was completely blocked (Fig. 5C). Taken together with these results and previous reports, we suggest that amitriptyline could block the 5-HT3 receptor before channel opening via binding to the closed ion channel.
Effects of amitriptyline on receptor desensitization
To find out whether amitriptyline could accelerate desensitization of 5-HT3 receptor or not, we compared the currents of 5-HT alone and 5-HT with amitriptyline for 15 second application (Fig. 6A), which protocol was already used to study desensitization [27]. In Fig. 6B, the desensitization time constant was decreased from 3838±1972 ms at 5-HT alone to 1304±506 ms at co-application of amitriptyline and 5-HT, and it was statistically significant (paired t-test, p<0.01, n=9). Decreased desensitization time constant at the 15 s long application could be understood that amitriptyline forces an opened 5-HT3 receptor to the desensitization, so it acts as an open channel blocker.
Simulation
To determine which step of receptor kinetic process amitriptyline acts on, a reaction scheme of agonist, receptor and antagonist were formulated (Fig. 7A). Binding of the agonist and channel, channel opening, channel closing, and channel desensitization were included in the reaction scheme. All reaction rate constants for each reaction steps were calculated and used to simulate macroscopic currents. Past studies used two or five binding of agonists to 5-HT3 receptor in the kinetic model [282930], however in this study, we formulated an one agonist binding model to estimate each rate constant since we simply supposed to which step was the target of the amitriptyline among the various channel states (close state, channel opening, and open state). Rate constants were calculated by the Berkeley Madonna® software and its results were shown in Table 1. The kf and kr were association and dissociation rate constants, respectively. The β and α were channel opening and closing rate constant, and the kd+ and kd− were desensitization and re-open from desensitization rate constant, respectively. Because we intended to elucidate the drug target based on current rising, decay and channel opening steps, we just compared kf, β, kd+, respectively. In Fig. 7B, the simulated currents generated by 5-HT alone, co-application and pre-application with amitriptyline were compared with experimental data. Simulated currents were closely overlapped with recorded the whole cell macroscopic data. The kf was significantly decreased by pre-application of the amitriptyline than other application mode (p<0.001, ANOVA and Tukey's multiple comparisons test, n=6). These results were matched to our macroscopic current data because kf were used to generate the rising slope (Table 1). So, currents slowly increased to the end of the 5-HT application in the pre-application mode. The β was not significantly changed by amitriptyline even in both application modes (p=0.89 by ANOVA, n=6), while kd+ was greatly increased in a co-application manner (p<0.001, ANOVA and Tukey's multiple comparisons test, n=6). From this study, we could expect that amitriptyline has less effect on the channel conformational change from a close to an open. And it is effective to force the channel to stay in the close state or change the channel state from open to close by accelerating 5-HT3 receptor desensitization.
DISCUSSION
In our study, amitriptyline inhibited the peak amplitude of 5-HT3 receptor current in a competitive manner and accelerated receptor desensitization. Its inhibitory effect was voltage independent and the currents were blocked by both pre-application and co-application modes.
By the pre-application of amitriptyline, currents evoked by 3 µM of 5-HT, which rapid rise and then fast decay, were changed to be similar with the currents induced by a low concentration of 5-HT, which the currents rose slowly to the end of 5-HT application (Fig. 5). However, in the co-application, the peak currents were decreased with the fast rising and fast desensitization. The inhibitory effect by the pre-application of a drug usually represents the close state blocking mechanism [2527]. In contrast, the inhibitory effect by the co-application or the use dependency represents the open state blocking mechanism of a drug [26]. If both of co-treatment and pre-treatment show inhibitory effect, the antagonist inhibits an ion channel via both of close state and open state blocking mechanisms [31]. Therefore our results suggest that amitriptyline acts as a close and open state ion channel blocker to the 5-HT3 receptor. Also we showed that the amitriptyline accelerated the receptor desensitization, which could support amitriptyline bound to the open state of ion channel and then forced the channel toward a desensitized state. Although voltage-dependent blocking of the ion channel is strong evidence of an open channel blocker, not all open channel blockers show a voltage dependence [32]. Because voltage dependency is properly related with a channel pore blocking mechanism, if antagonist binds at the outside of opened pore, it could show voltage independency. Taken together, we could suggest that amitriptyline acts as an open state blocker via binding at the outside of opened pore, and this binding site closely connected to the 5-HT binding on the 5-HT3 receptor, and act as a competitive blocker.
To compare each rate constant of recorded data when treating 5-HT alone, co-application with amitriptyline, and pre-application of the amitriptyline, the association rate constant kf, opening constant β, and desensitization constant kd+ were estimated by using the simulation (see Materials and Methods). Because the current trace was rising rapidly at the both of co-treatment with amitriptyline and 5-HT alone, there was no significant difference between the kf values. However, in the pre-application of amitriptyline, kf is significantly lower than 5-HT alone or with co-application. This current change by pre-application of amitriptyline is reasonable because 5-HT could force the dissociation of bounded amitriptyline from the receptor through a competitive manner, thus antagonist free 5-HT3 receptors are increased during the 5-HT application and generate slow rising inward current. The change of opening rate constant β caused by amitriptyline co- and pre-applications was statistically insignificant (Table 1). However, the desensitization rate constant kd+ was increased especially in the co-application method. The increase in kd+ due to amitriptyline in simulated data was consistent with our macroscopic current data, which showed desensitization was accelerated by amitriptyline only in the co-application mode (Fig. 6). In the pre-application mode, receptor desensitization might be occurred not much because current rise slowly and did not decay to the end of 5-HT application, thus kd+ was not significantly changed (Table 1). Although a macroscopic current recording technique useful to study the change of channel kinetics by a drug, it is still limited because it could not clarify the drug targeted-reaction step in the ion channel kinetics. Thus, a simulation study has frequently been used to interpret the ion channel kinetics change in combination with the macroscopic current recording [30]. In this study, we suggest that amitriptyline did not affect the channel opening conformational change in simulation because β was not changed greatly (Table 1). This result could not be proven by the analysis of macroscopic current. So we expect that our approach is useful to interpret a drug action mechanism on the 5-HT3 receptor.
Amitriptyline has been also used to treat migraine, fibromyalgia, neuropathic pain, and irritable bowel syndrome [151617]. However in the dorsal horn (the first synaptic site in nociception), an analgesic mechanism of amitriptyline is still unclear. From our study, we would suggest that 5-HT3 receptor is a potential target of the amitriptyline. 5-HT3 receptor is distributed at the dorsal horn of the spinal cord [33], connecting to the descending tracts from raphe-spinal neuron for the modulation of nociception [34]. However its pathophysiologic role in these chronic pain syndromes is still controversial. 5-HT3 receptor antagonist is known to block the GABA release at the dorsal horn, thus lack of GABA signals contributes a hypersensitivity of the peripheral nerve injury [35]. In contrast to this pro-nociceptive effect of the 5-HT3 receptor antagonist, the intra-thecal injection of ondansetron showed an analgesic effect in a spinal cord injury model, while agonist enhanced allodynia in the same experiment [36]. These results were interpreted that the 5-HT3 receptor regulated a pronociceptive neurotransmitter level at the synapse of dorsal horn [37]. This interpretation was also supported by the reports that 5-HT3 receptor activated the primary afferent fibers, including A-delta and C fiber in a patch clamp study [38], and 5-HT3 receptors is involved in the behavioral hypersensitivity activating the neuro-inflammatory signaling cascade [39]. Since amitriptyline is commonly used for the treatment of certain types of neuropathic pain, including diabetic neuropathy, post-herpetic neuralgia and other neuropathic pain [1640], our results, showing an inhibitory effect of amitriptyline on the 5-HT3 receptors, could be helpful to interpret the previous studies exploring the pathophysiologic role of 5-HT3 receptor in the neuropathic pain [1819].
Inhibition of the 5-HT3 receptor was expected to be related an antidepressive mechanism via increased monoamine contents in synapses, increased availability of other 5-HT receptor, and reducing inhibitory neurotransmission [1561241]. To understand the pathophysiologic role of 5-HT3 receptor in the depressive disorders, the effect of antidepressants on the 5-HT3 receptor and the effects of 5-HT3 receptor antagonist on depression models have been studied in various ways. Functional antagonism of the antidepressants on 5-HT3 receptor was reported in previous studies [212742]. Also ondansetron was known to improve a mood disturbance in the depressive patients and reduce anxiety behaviors in the mice models of depression [41434445]. Although these reports suggest the important role of 5-HT3 receptor in the depressive disorders, few antidepressants have been studied to test whether they modulated this receptor directly. Therefore we studied the direct effect of amitriptyline, even a classic tricyclic antidepressant, on the 5-HT3 receptor to expand our understanding the pathophysiology of depressive disorders.
In summary, we showed that amitriptyline directly inhibits the 5-HT3 receptor in close and open states ion channel in a competitive blocking mechanism. We could expand our understanding about the pharmacological mechanisms of amitriptyline in the treatment of depression and neuropathic pain, and the pathophysiologic role of 5-HT3 receptor, a potential target for a wide variety of neurologic and psychiatric diseases.




 XML Download
XML Download