

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Alopecia (hair loss) occurs due to many internal factors, such as aging or hormonal balance disruption, as well as diverse diseases and serious burns or wounds [12]. Hair loss is commonly accompanied by a region of thinned skin at the bald site [34]. Furthermore, these symptoms are due to the functional loss of follicular stem cell activity (i.e., forming the hair follicles) [5]. Recently, there have been numerous therapeutic trials of various treatments for hair loss such as oral medications and hair transplantation [67]. However, therapeutic drugs, like finasteride and dutasteride, provide only temporary improvement, and often with reduced effectiveness, which results in immediate hair loss [8910]. Although hair transplantation can replace the bald area with healthy hair, it is an invasive surgery, and a sufficient number of hairs for replacement is not always available in patients with severe hair loss [11]. Therefore, alternative therapeutic strategies are essential for alopecia.
The hair follicle (HF) cycle includes the phases of growth (anagen), regression (catagen), and rest (telogen) [12]. The activity of HFs can be influenced by the internal and external environments [13]. The mature HF has a complex structure with multilayered, concentric epithelial basement cylinders of keratinocytes and a distinctive mesenchyme of human dermal papilla cells (hDPCs) [14]. hDPCs are the main component of HFs and are widely studied as the key functional center responsible for controlling the hair cycle throughout an animal's life cycle [15]. The androgen activators dihydrotestosterone and dickkopf-1, which can initiate damage-induced apoptosis of hDPCs, are some of the many factors involved in androgenic alopecia, where the HF cycle is incrementally arrested during the telogen phase and in which scalp hair follicles are gradually replaced by smaller follicles [1617]. Thus, hDPCs may play a pivotal role in the pathogenesis of alopecia.
Human umbilical cord blood-derived mesenchymal stem cells (hUCB-MSCs) have been suggested to promote tissue repair and have been used for tissue engineering in numerous studies [1819]. Not only can their multiple lineages replace aged or damaged cells, but their secretory factors can enhance tissue repair efficiency [20]. Human mesenchymal stem cells (hMSCs), which are capable of self-renewal and differentiation into various mesenchymal tissues, including osteogenic, adipogenic, and chondrogenic cell lineages, have been recognized as a promising tool for clinical applications [21]. However, the secretion of soluble factors is one of the key mechanisms underlying the therapeutic potential of hUCB-MSCs recently shown to have therapeutic efficacy in various disease models [2223]. Transplantation of ex vivo-expanded hUCB-MSCs improved peripheral neuropathic pain [24] and focal cerebral ischemia [25] in rats. Thus, the treatment model of hUCB-MSC transplantation has been intensively studied for currently-untreated tissue impairments [26].
Hair loss from an adult follicle is considered to be a permanent state [2728]. However, there are many therapeutic approaches to overcome this irreversible outcome [29]. A previous study has demonstrated that HF neo-genesis can be induced in wounds in mice in an mIGF-1-dependent manner [30]. On other hand, hair follicle stem cell differentiation is disturbed via inhibition of Wnt/b-catenin [31]. The IGF-1/AKT/GSK3β/β-catenin signaling pathway is crucial for cell protection and tissue regeneration [3233]. The use of IGF-1/AKT/GSK3β/β-catenin on hDPCs in hair regeneration is groundbreaking. In this study, we used hUCB-MSCs to determine the effects of both stem cells and their paracrine factors on reacquisition of hDPC conduction ability to induce hair growth and to provide an effective therapeutic strategy for alopecia.
METHODS
Animals
C3H/HeJ mice were obtained from Saeron Bio Inc. (Gyeonggi-do, Korea) and allowed to adapt for one week with free access to food and water. The telogen-anagen transition model was induced by depilating the dorsal skin of mice in the telogen phase of the hair cycle, as described previously [34]. Starting one day after hair removal, minoxidill (3% v/w; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was topically applied to the minoxidill group (MNX) for five days per week. At one day after hair removal, 400 µl of saline or hUCB-MSCs (1×105 cells/mice) were intradermally injected into multiple points (8 sites) in the skin of the injection control and hUCB-MSC groups, respectively. The dorsal skin was harvested for histological analysis at the end of the observation period (six weeks). The skin was then fixed in 4% formaldehyde and processed by paraffin block embedding using standard techniques to produce paraffin sections (10 µm thick). The general histology was visualized using hematoxylin-eosin (HE) staining. Immunohistochemistry was performed on sections that had been steamed in 10 mM sodium citrate (pH 6.0, cat. no. 005000, Fisher Scientific, Inc., Waltham, MA, USA) for 15 min prior to incubation with primary antibody. We used antibodies directed against β-catenin (1:500, cat. no. 610153, BD Biosciences, Franklin Lakes, NJ, USA). Primary antibodies were detected by biotinylated secondary antibodies plus streptavidin-peroxidase complex (cat. no. SA-5004, VECTOR LABORATORIES, INC., Burlingame, CA, USA) and brown FAST DAB staining (cat. no. 34002, Thermo Fisher Scientific, Inc., Waltham, MA, USA). Counterstaining of sections was performed using hematoxylin for DAB-stained sections light microscopy (DM750, Leica, Wetzlar, Germany). The protocol was approved by the medical ethics committee of Chung-Ang University (Seoul, Korea).
Isolation and culture of hDPCs
hDPCs were purchased from Cefobio (Seoul, Korea) as primary cells and were grown in Dulbecco's modified Eagle's medium (DMEM; Invitrogen/Gibco-BRL, Grand Island, NY, USA) supplemented with 5% fetal bovine serum (FBS; Invitrogen/Gibco-BRL) and 1% penicillin in a humidified environment. hDPCs in the third or fourth passage were used.
Patch assay
The hair-inductive capacity of the hDPCs was assessed in accordance with the established patch assay [35]. Implantation was performed as described previously [36]. Briefly, hUCB-MSCs (U) were combined with freshly isolated neonatal mouse epidermal (K) and dermal cells (D) and co-transplanted subcutaneously into the skin on the backs of nude mice. The normal group, K plus D group, K plus U, and K plus D plus U group were injected on the same day. Two weeks later, skin samples were dissected from the mice and examined to verify hair induction.
Culture of hUCB-MSCs
This study was approved by the Institutional Review Board of MEDIPOST Co., Ltd. Neonatal hUCB-MSCs were collected from umbilical veins. Mononuclear cells were isolated from hUCB by centrifugation on a Ficoll-Hypaque gradient (density: 1.077 g/cm3; Sigma-Aldrich). The cells were then seeded at 5×105 cells/cm2 in culture flasks. After colonies of spindle-shaped cells formed, the cells were reseeded for expansion. hUCB-MSCs were cultured in αMEM medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10% FBS (Gibco) and gentamycin (Gibco) in a humidified atmosphere containing 5% CO2 and 3% O2 at 37℃. Cells were passaged when they reached 80% confluency and either used for experiments or redistributed to new culture plates. In all experiments, hUCB-MSCs used were at passage 6.
Human dermal papilla cell isolation and co-culture with hUCB-MSCs
For co-culture experiments, 1×105 hDPCs were seeded into the lower chamber of a 6-well transwell plate (Corning, New York, NY, USA) containing co-culture medium (DMEM or MEM). Immediately after the dermal papilla cells attached, the medium was changed, and 2×105 hUCB-MSCs were placed into the upper chamber. The plates were incubated at 37℃ with 5% CO2 for the indicated times before each analysis.
Cell proliferation assay
hDPCs were plated at a density of 1×105 cells/well in 6-well plates and were continuously cultured for 120 h under each test condition. At days 2 and 5 of co-culture with hUCB-MSCs, the proliferation was measured using a WST-8 assay (Dojindo; Rockville, MD, USA). WST-8 solution (100 µL) was added to the cells in 1 ml incomplete DMEM and incubated for 1 h at 37℃. The absorbance was then measured at 450 nm using a SpectraMax 190 microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Alkaline phosphatase (ALP) activity
hDPCs were seeded at a density of 1×105 cells/mL in 6-well plates. At days 2 and 5 of co-culture with hUCB-MSCs, ALP activity of the hDPCs was determined enzymologically using an ALP assay. After washing with 1X PBS, the cells were incubated in 0.1 M NaNO3-Na2CO3 (Sigma-Aldrich) buffer containing 1% (v/v) Triton X-100 (Sigma-Aldrich) and 2 mM MgSO4 (Sigma-Aldrich). Subsequently, 6 mM p-nitrophenyl phosphate (Sigma-Aldrich) was added as a substrate to each well and incubated for 30 minutes at 37℃. Finally, 1.5 M NaOH (Sigma-Aldrich) was added to stop the enzyme-substrate reaction. Optical density (OD) readings were performed at a wavelength of 405 nm using a SpectraMax 190 microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Human growth factor and cytokine antibody array
The Raybio® Human Cytokine/Growth Factor Antibody array I (RayBiotech, Norcross, GA, USA) was used to assay 23 cytokines and 41 growth factors in the supernatants of the cell cultures. The array membranes were incubated in blocking buffer for 30 min at room temperature (RT), then 1 ml of conditioned medium from either the hDPC, hUCB-MSCs, or hDPC plus hUCB-MSCs group was added per well, followed by incubation for 1 h at RT. The membranes were washed five times in wash buffer at RT, a biotin-conjugated antibody was added, and the membranes were incubated for 1–2 h at RT. The membranes were washed again, 2 ml of HRP-conjugated streptavidin was added, and the membranes were incubated for 2 h. Following incubation with detection buffer for 2 min, the luminescence was detected using the LAS 3000 chemiluminescence imaging system (Fujifilm Inc., Tokyo, Japan) and analyzed by densitometry using ImageJ 1.44 software (National Institutes of Health, Bethesda, MD, USA).
ELISA
The levels of insulin-like growth factor-1 (cat. no. DG100, R&D Systems, Minneapolis, MN, USA), insulin-like growth factor binding protein-1 (cat. no. DGB100, R&D Systems), and vascular endothelial growth factor (cat. no. DVE00, R&D Systems) in cell culture supernatants were quantified using an immunoassay kit. After each experiment, the collected medium was added to each well and incubated at room temperature for 2 h. The protein expression level was determined according to the manufacturer's protocol, and absorbance was measured at 450 nm using a SpectraMax 190 microplate reader (Molecular Devices).
Western blot analysis
Western blotting was performed as previously described using total protein obtained from hDPCs. Antibodies for phosphorylated-AKT (1:1000, cat. no. #9271, Cell Signaling Technology, Inc., Danvers, MA, USA), AKT (1:1000, cat. no. #4685, Cell Signaling Technology, Inc.), phopholyrated-GSK3β (1:1000, cat. no. #9323, Cell Signaling Technology, Inc), GSK3β (1:1000, cat. no. #12456, Cell Signaling Technology, Inc.), β-catenin (1:1000, cat. no. 610153, BD Biosciences, Franklin Lakes, NJ, USA), proliferating cell nuclear antigen (1:1000, cat. no. #13110, Cell Signaling Technology, Inc.), β-actin (1:1000, cat. no. sc-4778, Santa Cruz Biotechnology, Inc., Dallas, TX, USA), ALP (1:100, cat. no. SC-15065, Santa Cruz), CD 133 (cat. no. NB120-16518, Novus Biologicals, CO, USA) were used as the primary antibodies, and goat anti-rabbit (cat. no. BA-1000, VECTOR LABORATORIES, INC.) and goat anti-mouse (HRP) (cat. no. BA-9200, VECTOR LABORATORIES, INC.) were used as the secondary antibodies (1:10000). The blots were analyzed using densitometry with ImageJ 1.44 software (National Institutes of Health).
Establishment of hDPC micro-tissues in 3D hanging drop cultures
For 3D cell cultures, Perfecta3D Hanging Drop Plates (3D Biomatrix Inc., Ann Arbor, MI, USA) were used according to the manufacturer's protocol. To test the effects of IGFBP-1 on hDPCs micro-tissue formation, the rhIGFBP-1 (cat. no. 871-B1-025, R&D Systems) (+) group was treated with rhIGFBP-1 (100 ng/ml). The size distribution of the resulting hDPCs micro tissues was analyzed to determine the optimal culture conditions. All hDPCs were cultured in serum-free medium (DMEM), which was changed every 2 days using a multichannel pipette. The morphology and diameter of the hDPC spheroids were recorded using a reverse phase-contrast microscope (IX61 FL, Olympus Japan Co.) for up to 5 days.
Immunofluorescence staining
Immunofluorescence staining in hDPCs was performed as previously described [37]. Briefly, hDPCs were treated with rhIGFBP-1 (100 ng/ml) for 2 days. The cells were fixed in 10% neutral buffered formalin solution at room temperature for 30 minutes, permeabilized in 2.5% Tween 20 (cat. no. 9005-64-5, Sigma-Aldrich) in PBS for 5 min, then incubated overnight at 4℃ with anti-ALP (1:500, cat. no. SC-15065, Santa Cruz), anti-β-catenin (1:500, cat. no. BD610154, BD Biosciences), anti-CD 133 (cat. no. NB120-16518, Novus Biologicals, CO, USA), anti-IGF-1 (cat. no. SC-9013 Santa Cruz), or anti-IGFBP-1 (cat. no. sc25257, Santa Cruz) primary antibody in blocking solution (1:1000). After several washes with PBS containing 0.5% Triton X-100 and 0.5% BSA, the hDPCs were incubated with anti-rabbit Cy2 (cat. no. 711-225-152, Jackson ImmunoResearch Inc., PA, USA) or anti-mouse (cat. no. 715-165-150; Jackson ImmunoResearch Inc.) labeled secondary antibodies for 1 h at RT. Following nuclear staining with DAPI dye (Sigma-Aldrich), the fluorescence signals were viewed using a Zeiss confocal laser microscope (LSM 700; Zeiss, Heidelberg, Germany).
RESULTS
hUCB-MSCs promote hair follicle cycling, morphogenesis in vivo
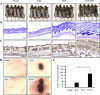
To determine whether hUCB-MSCs promote the anagen hair cycle, we injected hUCB-MSCs intra-dermally at multiple sites in depilated mouse dorsal skin. Saline injection and daily topical treatments of 3% minoxidil were used as controls. We determined the hair cycle stage by measuring the status of skin hair regrowth. We found that, by 5 weeks of treatment, hUCB-MSCs caused diffuse darkening of the dorsal skin, while the control groups showed no significant changes (data not shown). At 6 weeks of treatment, hair regrowth was complete in the hUCB-MSCs group, and the tip of the hair shaft emerged through the epidermis. Although the dorsal skin in the minoxidil-treated group had incomplete pigmentation and contained hairs in the early stage of the hair cycle, the dorsal skin of the normal and saline-injected groups retained large areas without anagen induction (Fig. 1A). After 6 weeks of treatment, we harvested the dorsal skin for histological analysis, which indicated that the hUCB-MSCs promoted the telogen-anagen transition (Fig. 1B). In particular, the hair follicles that were treated with hUCB-MSCs were transformed from the telogen phase to the early- and middle-anagen phases at 6 weeks (Fig. 1B). Remarkably, the hUCB-MSCs group prematurely reentered the anagen phase compared with the minoxidil-treated group (Fig. 1B). Furthermore, IHC data revealed that expression of the β-catenin protein, which is a positive regulator of hair growth, was up-regulated in the hUCB-MSCs group compared to the other groups (Fig. 1C).
To verify how hUCB-MSCs affect hair follicle morphogenesis, we implanted hUCB-MSCs (U) with epithelial stem cells (K) and dermal stem cells (D) of neonatal mice into nude mouse. Mixed U, K, and D exhibited increased hair follicle induction compared to the K plus D group (Figs. 1D and E). Interestingly, the K plus U group did not show hair follicle morphogenesis (Figs. 1D and E). Collectively, hUCB-MSCs enhanced hair follicle morphogenesis, represented as an increase of follicular neogenesis (Figs. 1D and E).
hUCB-MSCs increase cell viability and upregulate hair induction-related proteins in hDPCs
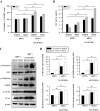
Changes in the microbiological environment cause hDPCs to lose their conduction ability to induce hair follicle formation in humans. To study the effects of hUCB-MSCs on the viability and characteristics of hDPCs, we estimated cell viability and ALP activity. The effect of hUCB-MSCs on hDPC proliferation and ALP activity was revealed using WST-8 and ALP-activity assays. hUCB-MSCs co-culture enhanced hDPC proliferation (Fig. 2A) and restored ALP activity, both of which are anagen markers of the hair cycle in hDPCs in vitro (Fig. 2B) [38].
We further examined AKT/GSK3β/β-catenin pathway regulation and proliferating cell nuclear antigen (PCNA) expression. Result revealed that protein expression of phosphorylated Akt (Ser473), phosphorylated GSK3β (Ser9), β-catenin, and PCNA was pronounced in the hDPCs plus hUCB-MSCs group compared to hDPCs only. These results indicate that β-catenin, AKT, and GSK3β, which are proteins contained in the pathway related to cell growth and proliferation, were up-regulated in hDPCs by co-culture with hUCB-MSCs. We found that hUCB-MSCs not only promoted expression of hair anagen-induction-related proteins, but also upregulated the expression of PCNA in hDPCs (Figs. 2C and D). These findings suggest that, in hDPCs, the viability and ALP activity related to anagen conduction ability can be affected and repaired by hUCB-MSCs in vitro, and this effect is mainly regulated by AKT/GSK3β/β-catenin signaling.
Growth factor production in hDPC cultures with hUCB-MSCs
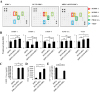
A screening of growth factor production was performed to discover the mechanisms underlying hair anagen induction by hUCB-MSCs. Culture media supernatants from hDPCs, hUCB-MSCs, and hDPCs plus hUCB-MSCs were collected after 5 days of culture and analyzed with an antibody array for 42 growth factors. Although several factors were detected in the hDPC plus hUCB-MSCs group, two growth factors were upregulated in the medium of the hDPC plus hUCB-MSCs group compared to the hDPC group (Figs. 3A and B). The hDPCs co-cultured in the presence of hUCB-MSCs showed a considerably increased levels of IGFBP-1 (about >1.25-fold) and VEGF (about >1.25-fold). Notably, IGFBP-1 was detected in the hUCB-MSCs-only group but less in the hDPC-only group. VEGF was also detected in the hUCB-MSCs groups, but there was no significant difference compared to the hDPC-only group. In addition, secretion of IGFBP-2, -6, and PDGF-AA were detected both in the hDPCs and hDPCs plus hUCB-MSCs group, but there was no statistical significance to the increase. In conclusion, it was confirmed that the secretory source of IGFBP-1 is hUCB-MSCs.
The actions of IGFs are tightly modulated by a family of proteins called IGF-binding proteins (IGFBPs), of which IGFBP-1 to -6 have been characterized [39]. We measured IGF-1 and IGFBP-1 concentrations in culture media with an ELISA assay. Consistently, IGFBP-1 was not detected in the hDPCs alone, but the hUCB-MSCs and hDPC plus hUCB-MSCs group showed increased IGFBP-1 level compared to hDPCs alone (Fig. 3C). Additionally, VEGF concentration was detected in the hDPC alone group and hUCB-MSCs alone group, but it was overwhelmingly upward in the hDPC plus hUCB-MSCs compared to each alone group (Fig. 3D). These results suggest that the IGFBP-1 expression measured by the growth factor array is primarily secreted by the hUCB-MSCs, and this is related to VEGF concentration. In addition, we measured IGF-1 concentration in the culture media to determine the relationship between IGF-1 and IGFBP-1 expression. The protein concentration of IGF-1 was decreased in the hDPC plus hUCB-MSCs group compared to the hDPC group (Fig. 3E).
IGFBP-1 affects dermal papilla cell viability and promotes expression of diverse proteins related to hair induction
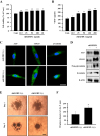
IGFBP-1, which was confirmed as the main component secreted by hUCB-MSCs, was studied to explore its independent function. To investigate the effects of IGFBP-1 on hDPCs, we treated hDPCs with recombinant human IGFBP-1 (rhIGFBP-1) protein and measured cell viability and VEGF secretion. The results indicated that the viability of hDPCs was significantly increased at 48 h after rhIGFBP-1 treatment (rhIGFBP-1 concentration: 12.5–200 ng/ml) (Fig. 4A). Furthermore, VEGF secretion also increased in an rhIGFBP-1 concentration-dependent manner in hDPCs (Fig. 4B). The expression of proteins related to hDPC conduction ability was confirmed by immunofluorescence in hDPCs. ALP and CD133 staining were strong after rhIGFBP-1 treatment (100 ng/ml) compared to the non-treatment group (rhIGFBP-1 (−)). Additionally, β-catenin expression and nuclear translocation were increased in the rhIGFBP-1 (+) group (Fig. 4C). Western blot data also revealed the upregulated protein expression of ALP, CD133, non-phosphorylation-β-catenin (non-p-β-catenin, activation form), and β-catenin in the rhIGFBP-1 (+) group (Fig. 4D). To investigate the characteristics of hDPCs regarding the transformation to spheres and hair induction structures, the 3D sphere formation velocity of hDPCs was tested by seeding them in hanging-drop 96-well plates. When the seeding density exceeded 0.25×104 cells in the hanging-drop 96-well plate, the rapid aggregation of hDPC with a loose spheroid structure occurred on day 1, which then changed to a compact circular sphere after 5 days of incubation (Fig. 4E). The diameters of the spheres were quantified using phase contrast imaging on day 5. After seeding the hDPCs in hanging-drop 96-well plates, hDPCs treated with rhIGFBP-1 (100 ng/mL) produced spheres with an approximate diameter of 200 µm, which was larger than the sizes in the rhIGFBP-1 (−) group, at day 5. The maximum diameter of the rhIGFBP-1 (−) group was approximately half that of the rhIGFBP-1 (+) group (Figs. 4E and F). Collectively, these data support the idea that rhIGFBP-1 can stimulate cell proliferation; increase secretion of VEGF and enhance expression of ALP, CD133, and β-catenin; and promote the conduction ability to 3D tissue-like formation.
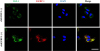
Effects of recombinant protein IGFBP-1 on co-localization of IGF-1 to dermal papilla cells
In previous reports, it was demonstrated that IGFBP-1 can act either directly or indirectly in biological environments [40] and form the complexes with IGF-1 [41]. We hypothesized that increased IGFBP-1 acts as a complex with IGF-1 in hDPCs. Therefore, we investigated the effects of rhIGFBP-1 treatment on the expression of IGF-1 in hDPCs. The results showed that most IGF-1 was co-localized with IGFBP-1, which was assumed to be the main secretory protein of hUCB-MSCs (Fig. 5). Interestingly, the protein expression of IGF-1 was slightly elevated in the rhIGFBP-1 (+) group compared to the rhIGFBP-1 (−) group (Fig. 5). These results suggest that secreted IGFBP-1 in hUCB-MSCs co-locate with IGF-1 and affects the expression of IGF-1 in hDPCs.
DISCUSSION
Alopecia is a serious dermatological disease that occurs due to diverse reasons [42]. Genetic factors and age have typically been considered the main causes of hair loss [43]. Androgenic alopecia results in decreased viability of dermal papilla cells and other cells comprising hair follicle tissue [16]. Dermal papilla cells play a central role in follicular growth and differentiation and direct hair follicle cycling by secreting signaling factors to communicate to root sheath cells [44]. The hair follicle cycle is affected by androgen and several other factors produced by stressful environments, which cause a primary defect in hDPCs signaling, resulting in an inability to initiate the hair follicular anagen phase [4546]. Therefore, the restoration of hDPCs hair induction ability has been considered a potential therapy for hair loss.
Mesenchymal stem cells (MSCs) exist as a heterogeneous population in animals and have great potential for use in tissue regeneration [47]. These MSCs are usually distinguished by their shape, small size, and high nuclear fluctuation, which determine their “stemness” and therapeutic capacity [48]. Attempts to regenerate tissue using biological factors secreted by UCB-MSCs have recently been reported [4950]. In a previous study, implantation of BM-MSCs in normal and diabetic mice enhanced wound healing associated with increased angiogenesis [51]. In another study, mesenchymal stem cell transplantation has beneficial results due to its anti-inflammation potential in cardiac infarction damage [52]. In addition, intravenous injection of hUCB-MSCs has a role in the prevention of obesity as a mediator in macrophage polarization [53]. The effects of MSCs on tissue regeneration and anti-inflammatory effects have been verified, and the functions and main factors causing the therapeutic effects of MSCs have been widely explored [54]. In this study, hUCB-MSCs triggered hair anagen induction from the telogen phase in vivo. Furthermore, hUCB-MSCs emphasized hair follicle morphogenesis, but in case of ablation of the neonatal dermal cell, hUCB-MSCs and epithelial cells did not undergo hair follicle generation. We assumed that hUCB-MSCs are not the main component of the newly generated hair follicle and may play a role in assisting dermal cells to conduct tissue generation. These findings indicate that hUCB-MSCs facilitate hair follicle regeneration and hair growth via a paracrine mechanism. In a previous report, MSC-conditioned medium that was modified to overexpress Wnt1a restored the hair cycle in vivo [34]. In another study, BM-MSCs increased angiogenesis in a diabetic mice wound model [51]. However, BM-MSCs were located adjacent to the vasculature rather than in the vascular walls [51]. This indicates that a paracrine effect of MSCs plays a major role in angiogenesis and wound healing. In this study, we demonstrated that hDPCs co-cultured with hUCB-MSCs in a detached environment had increased cell viability and ALP activity and up-regulated the AKT/GSK3β/β-catenin pathway, indicating that paracrine factors have a major role in recruitments of hDPC conduction ability in vitro.
Insulin-like growth factor 1 (IGF-1) is an important growth factor in many biological environments [5556]. In a previous report, it was found to have substantial structural homology with insulin and influence food intake and glucose metabolism in a similar manner to insulin [57]. Recently, IGF-1 has been suggested to play a crucial role in regulating cellular differentiation and tissue generation during the development of hair follicles [58]. To exert its physiological effects, IGF-1 activates cells by binding to specific cell-surface receptors [58]. In the circulatory system, this biological factor mediates the endocrine action of growth hormones on cellular growth and forms complexes with specific binding proteins (BPs) [41].
There are six insulin-like growth factor binding proteins (IGFBP-1~-6), which bind to IGF-1 and -2 with very high affinity [41]. Because their affinity to IGFBP is 2- to 50-fold greater than that of IGF-IR, these IGFBPs can modulate the distribution of IGF among the proteins in interstitial fluids [41]. In this study, we hypothesized that IGFBP-1 was be a major factor affecting dermal papilla cells. We assumed that, since IGF-1 protein is the autocrine factor of hDPCs bound to hDPCs with the assistance of IGFBP-1, its concentration in culture media was decreased compared to that of the non-assisted group (hDPCs alone).
Cells within hDPCs and dermal sheath (DS) are specially identified as mesenchymal cells and express distinct enzymes and molecules. Although the functions of most of these protein markers are not fully understood, they have been used to identify hDPCs and DS. The expression of some markers, like ALP and versican, correlates with hair inductive properties [59]. hDPCs easily lose their characteristics and conductivity in vitro in the absence of additional factors, such as WNT- or BMP-signaling [60]. In a previous report, beta-catenin was upregulated in hDPCs, resulting in increased expression of the mesenchymal stem cell marker CD133 [61]. Furthermore, this anagen-conductive protein is also increased during the formation of hDPCs 3D tissue-like aggregates in vitro and in vivo [62]. In this study, rhIGFBP-1 not only increased hDPCs viability and VEGF secretion, but also upregulated the protein expression of ALP, beta-catenin, and CD133. In addition, rhIGFBP-1 enhanced the formation velocity of 3D tissue-like aggregates of hDPCs in vitro. These results indicate that IGFBP-1 has a positive role in maintaining cell longevity and regulating the conduction ability of hair anagen momentum.
IGFBPs can affect cells directly or indirectly and modulate the function of IGF-1 in an endocrine, paracrine, or autocrine manner through IGF-1/IGFBP complexes [41]. In a previous report, when IGFBP-1 and IGF-1 were added to porcine aortic smooth muscle or human fibroblast cultures, the IGF-1/IGFBP-1 complex enhanced DNA synthesis compared to treatment with IGF-1 alone [41]. Additionally, IGFBP-1 allowed IGF-1 to remain in equilibrium with its high-affinity receptors for a prolonged interval [4041]. In our results, rhIGFBP-1 formed a co-localization with IGF-1. Notably, this result suggests that the IGF-1/IGFBP-1 co-localization facilitates the interaction of IGF-1 with its receptor in hDPCs during the progression of the cell cycle.
In this study, we demonstrated that hUCB-MSCs can accelerate the initiation of the hair follicle telogen-anagen transition, increase the number of hairs in vivo, and enhance expression of proteins related to hair induction in vitro. Notably, IGFBP-1 (assumed as the main secretory factor of hUCB-MSCs) restores and promotes the hair-induction ability of hDPCs via an IGF-1/IGFBP-1 co-localization. Taken together, our findings indicate that hUCB-MSCs and their secretory protein IGFBP-1 can restore the ability of hDPCs to induce hair follicle regeneration, potentially providing alternative therapeutic methods for alopecia. In general, the immunosuppressive effect of the mesenchymal stem cell treatment is well known, as mentioned above. The occurrence of hair loss is also complex, and the immune response is one of its major causes. We would need to further investigate the effects of stem cell therapies on hair loss in a variety of biological environments.
 XML Download
XML Download