

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Japanese hop [Humulus japonicus (H. japonicus)] is a major cause of weed pollinosis in East Asia.1 Its pollen count and skin test positivity rates in Korea have increased significantly over the past 6 years.2 Analysis of 6043 subjects, mostly Han and Mongolian tribes, in northern China revealed that Japanese hop, along with mugwort and chenopodium, is the most frequent cause of allergic rhinitis.3 Moreover, high levels of IgE to Japanese hop or mugwort have been described as increasing the risk of asthma development in northern China.4 Furthermore, Japanese hop was listed as an invasive alien plant in Europe in 2007 and in the United States in 2012.
Development of allergy diagnostics and immunotherapeutics is largely dependent on supplies of qualified allergen extract. While Japanese hop pollen reference material is currently prepared using standardized methods,5 Japanese hop cultivation and allergen manufacturing has failed to meet clinical demand, and a limited supply of allergen extract has made allergy diagnosis and immunotherapy thereof difficult.
The content of pollen allergens is affected by environmental factors, such as weather, air pollution, and pathogens present during plant growth.6 Previously, ragweed pollen extract prepared using the same method with pollen collected from the same location in 1980 and 1995 yielded roughly 10-fold different quantities of the allergen, Amb a 1.7 Current commercial Japanese hop extract products are developed using pollens collected from wastelands. To reduce batch-to-batch variation, cultivating plants in a farm or greenhouse would be more desirable.
Common hop [Humulus lupulus (H. lupulus)] causes allergic disease in occupational settings, especially in the beer brewing industry.89 Since H. lupulus and H. japonicus are closely related species, cross-reactivity might be expected. In this study, we sought to examine IgE cross-reactivity between pollen extracts from these two hops to determine whether extracts from cultivated common hop could replace those from Japanese hop for development of diagnostic and immunotherapeutic reagents.
Go to : 
MATERIALS AND METHODS
Allergen extracts
Japanese hop pollen extract was obtained from Prolagen (Seoul, Korea), and lyophilized common hop pollen extract was obtained from Lofarma (Milano, Italy). Pollen extracts were reconstituted in phosphate-buffered saline (PBS) solution for Japanese hop and distilled water for common hop, aliquoted, and kept frozen before use.
Serum samples
Serum samples were collected from 7 patients (average age, 30 years; age range, 12–59 years) who visited the Allergy-Asthma Center at Severance Hospital, Seoul, Korea (Table 1). Allergy diagnosis was based on patient history and a skin prick test. Specific IgE reactivity to Japanese hop was determined using an ImmunoCAP assay (Thermo Fisher Scientific, Uppsala, Sweden). Methods used to collect serum samples were approved by the Institutional Review Board of our institution (4-2017-1197).
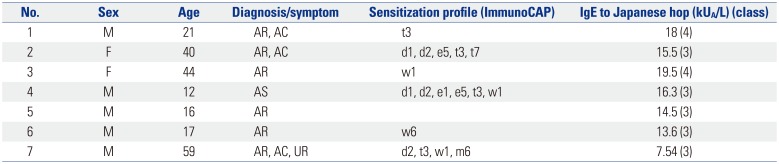
Table 1
Clinical Features of Enrolled Subjects
![]()
Inhibition analysis
Inhibition ELISA was performed to examine IgE cross-reactivity between Japanese and common hop pollen extracts as follows. Japanese hop pollen extract (10 µg/mL) was added to a microplate (Corning Inc., Corning, NY, USA). After blocking with 3% skim milk in PBS containing 0.05% Tween 20 (PBST), the plate was incubated overnight with 1:4-diluted pooled serum samples that were pre-incubated with various concentrations (0.00064–10 µg/mL) of pollen extract. Subsequently, IgE antibodies bound to Japanese hop were detected using biotinylated goat anti-human IgE (1:1000) (Vector, Burlingame, CA, USA) and peroxidase conjugated with streptavidin (1:1000) (Sigma-Aldrich, St. Louis, MO, USA). Color was developed using KPL SureBlueTM TMB Microwell Peroxidase Substrate (1-Component) (Kirkegaard & Perry Laboratories, Gaithersburg, MD, USA). The reaction was stopped by adding 0.5 M H2SO4, and absorbance at 450 nm was measured. Percent inhibition was calculated as (1-absorbance with inhibitors/absorbance without inhibitor)×100.
SDS-PAGE and IgE immunoblot
Pollen extracts (20 µg/well) were run on a 18% SDS-PAGE gel under reducing conditions and stained with Coomassie Brilliant Blue R-250 or transferred to a polyvinylidene difluoride (PVDF) membrane (GE Waters and Process Technologies, Trevose, PA, USA). After blocking with 3% skim milk in PBST, IgE-reactive components were probed overnight using 1:4-diluted pooled serum from the 7 patients. IgE-bound proteins were detected using alkaline phosphate-conjugated goat anti-human IgE (1:1000) (Sigma-Aldrich) for 1 h, and color was developed with 1-StepTM NBT/BCIP Substrate Solution (Thermo Fisher Scientific, Rockford, IL, USA).
Identification of IgE-reactive components
After desalting with trichloroacetic acid, two-dimensional (2D) gel electrophoresis was performed to analyze IgE-reactive components. 50 µg of each protein was separated on an 18% SDS-PAGE gel after isoelectric focusing (pH 3–10). Proteins in the gel were stained with Coomassie Brilliant Blue or transferred onto a PVDF membrane. IgE-reactive components were detected as described above. IgE-reactive proteins were excised, and in-gel tryptic digestion was performed. Protein identification was performed using liquid chromatography coupled to electrospray ionization tandem mass spectrometry (LC-ESI-MS/MS).
Go to :
RESULTS
IgE cross-reactivity between Japanese and common hop pollen extracts
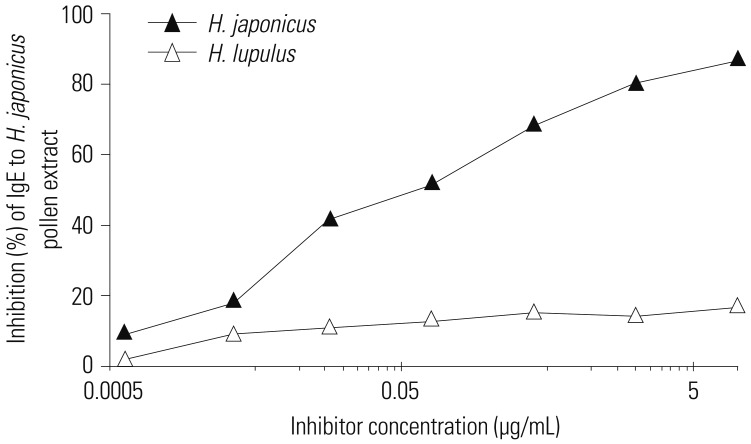
Inhibition of IgE reactivity to Japanese hop by common hop was examined by ELISA. IgE reaction to Japanese hop was inhibited up to 16.7% by common hop, while Japanese hop itself showed a maximum of 86.7% inhibition (Fig. 1).
IgE-reactive components of Japanese hop pollen
IgE-reactive components of each pollen extract were compared using one-dimensional (1D) and 2D gel electrophoresis combined with IgE immunoblotting. SDS-PAGE revealed large differences in the protein components of the two pollen extracts (Fig. 2). Specifically, there were more proteins in the 10–20 kDa range in Japanese hop. There were >10 IgE-reactive protein components in the 11–70 kDa range detected in Japanese hop, compared to only five in common hop. Notably, a 12 kDa protein component displayed the strongest IgE reactivity in Japanese hop, while no IgE reaction to a 12 kDa component was observed for common hop.

In a 2D gel, 11–20 kDa proteins were observed in Japanese hop, but not in common hop. Six spots exhibited IgE reactivity in Japanese hop, and none in common hop (Fig. 3).
 | Fig. 3Proteomic analysis of hop pollen proteins. Proteins (50 µg) were separated and visualized with Coomassie Blue stain, and IgE-reactive components were probed with serum from Japanese hop allergy patients. (A) Japanese hop protein profiles. (B) Common hop protein profiles. (C) IgE-reactive components of Japanese hop. (D) IgE-reactive components of common hop. Proteins indicated with arrows were excised and subjected to liquid chromatography coupled to electrospray ionization tandem mass spectrometry analysis.
|
LC-ESI-MS/MS was performed to identify the six IgE-reactive components (Table 2). However, the Mascot score for the six proteins (ranging from 59 to 70) was insufficient for positive identification.
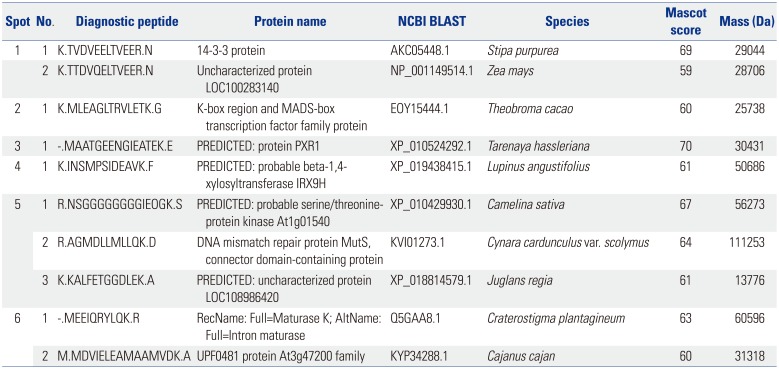
Table 2
Identification of IgE-Reactive Components from Japanese Hop Pollen
![]()
Go to :
DISCUSSION
High-quality raw material is essential for standardization of allergen extracts. This study was initiated to test whether common hop pollen, which can be obtained in large quantity from cultivated fields, can be used to replace Japanese hop pollen, a major cause of seasonal allergic rhinitis in East Asia, as a source of allergen extract for diagnostic and immunotherapeutic reagents. However, no significant IgE cross-reactivity was detected between Japanese and common hop pollen extracts (Fig. 1). The results are consistent with a recent study in which no significant cross-reactivity between these species was observed in Korean patients with Japanese hop pollinosis.10
The two pollen extracts showed different profiles of protein and IgE-reactive components based on IgE immunoblotting of sera from Korean patients with Japanese hop pollinosis (Fig. 2). In 1D gel analysis, at least 10 different IgE-reactive components were detected in Japanese hop, whereas only five components were observed in common hop. A 12-kDa component in Japanese hop exhibited strong IgE reactivity, but showed no counterpart in common hop. Interestingly, no IgE-reactive component of common hop was detected in 2D gel analysis. This discrepancy between the 1D and 2D gel results may be partly explained by primary sensitization to Japanese hop and the low avidity of Japanese hop-specific IgE antibodies to common hop protein. Relatively low concentrations of proteins in the 2D gel may also account for the lack of detection of IgE reaction. It is possible that a minor component of Japanese hop is cross-reactive with a common hop counterpart. However, it is clear that the 12 kDa component of Japanese hop is the major allergen and that this component is not cross-reactive with common hop allergens. Potent IgE reactivity to a 12 kDa allergen is consistent with our previous study.11 Furthermore, four patients were also sensitized to birch, two patients to ragweed, one patient to oak, and one patient to mugwort (Table 1). The multiple sensitizations of these patients to pollen allergens suggest possible IgE reactions to plant panallergens, such as Bet v 1 homologues, profilins, and calcium binding proteins.12
For identification of IgE-reactive components, LC-coupled ESI-MS/MS was performed. However, Mascot scores were insufficient for positive protein identification. No protein identified showed homology to previously described allergenic molecules. This may be due to poor database quality for this plant species. Construction of a high quality database for Japanese hop using expressed sequence tags or RNA-seq may be beneficial.
To date, profilin is the only Japanese hop protein that has been identified as an allergen.13 Previously, however, profilin was not found to be a major allergen in Korea,14 although it showed strong IgE reactivity to five sera from Chinese pollinosis patients.13 All five of these Chinese patients were also sensitized to ragweed allergen Amb a 8 (profilin), implying a cross-reaction between profilin allergens. The allergenicity of Japanese hop profilin was shown to increase following exposure to ozone and nitrogen dioxide, indicating the importance of environmental factors on pollinosis.15
In conclusion, there is no significant cross-reactivity between Japanese and common hop pollen allergens. Common hop pollen cannot be used to replace Japanese hop pollen for development of allergy diagnostic and immunotherapeutic reagents. Farm cultivation of Japanese hop is essential for allergen standardization. Furthermore, since the molecular identity of the major allergen from Japanese hop pollen remains unclear, characterization of this allergen is urgently needed.
Go to :
 XML Download
XML Download