

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Toxoplasmosis is a cosmopolitan zoonotic disease, caused by an intracellular protozoan belonging to the Apicomplexa phylum that has a worldwide distribution [123]. Over one third of the human population are chronically infected worldwide [456]. Cats are the only definitive hosts, and several warm-blooded mammals, including humans, rodents, birds, etc. serve as intermediate hosts [3789]. Additionally, Toxoplasma gondii was reported in snakes. Nasiri et al. (2016) [10] in Iran showed that 80.88% (55/68) of the examined snakes were positive by GRA6 gene and sequencing revealed over 98% similarity with T. gondii available sequences in GenBank [10].
A wide range of risk factors are involved in the prevalence of toxoplasmosis, so that they can affect the global/regional epidemiological figure of infection including: close contact with cats or keeping them indoors as pet animals, occupation, place of residence, education level, age, eating raw meat, gender, exposure to soil, the host immune response, etc. [45111213]. In humans, toxoplasmosis is often asymptomatic in immunocompetent persons, although in immunocompromised subjects such as patients with malignancies, human immunodeficiency virus-positive individuals, and organ transplant recipients may be cause severe and progressive complications with poor prognosis or even may result in death if not treated [5131415]. Besides, seronegative pregnant women are other risk groups for T. gondii infection [1617]. Upon maternal infection, fetus is probably to be exposed with transplacental transmission. Toxoplasmosis may cause miscarriage in those pregnant mothers that acquired the infection during their pregnancy [1819]. In general tachyzoites as an infectious form of parasite T. gondii are able to actively invade all nucleated cells of the intermediate host and their replication is ultimately curtailed by protective immune response [20].
The present common primary control measures for men and animals toxoplasmosis depends on chemotherapy. There are very few effective control strategies to limit infection and disease in humans and numerous warm-blooded animals throughout the globe and unfortunately the methods of therapy still could not fulfil entirely the treatment goals [21]. At the moment, the drugs for treatment of toxoplasmosis is a combination of pyrimethamine and sulfadiazine that has several side effects. Furthermore, these drugs are expensive and inadequate, which may result in toxic hypersensitivity reactions and are teratogenic on the fetus as well as they cannot eliminate bradyzoites into tissue cysts [21]. Considering the high prevalence of toxoplasmosis in the world, the resulting clinical, mental, and economical complications, as well as the current common drugs have no effect on the encysted parasites [2212223]; therefore, the development of a vaccine against T. gondii parasite can be important and necessary for preventing infection [2425262728].
In recent decades, numerous advancement has been made in order to identification of vaccine candidates against both chronic and acute toxoplasmosis that could promote an effective immune response. Hence, most of the work in the development of T. gondii vaccines have focused on dense granule antigens (GRA), microneme antigens (MIC), rhoptry antigens, surface antigens (SAG), and some other antigens. In this regard, a wide variety of vaccines such as DNA vaccines, recombinant protein vaccines, etc. have been investigated in many countries [24252627293031323334353637]. Despite of the constant efforts of scientists, there is no commercial vaccine for use in human and animals.
Microneme Proteins (MICs)
During recent decades, the increasing number of articles have focused on the evaluation of the immunogenicity of several functional proteins that involve in motility, adhesion to host cells, migration, invasion, and establishment of the parasitophorous vacuole [3839]. Among these, MICs secreted by the micronemes play a critical role in the initial stages of host cell invasion by parasites that are located at the apical end of the zoite and surrounded by a typical unit membrane [3840]. MICs are produced at the rough endoplasmic reticulum, then are transferred to micronemes by the Golgi apparatus in order to participate in cell attachment. In the other hands, these proteins are released by micronemes after contact between parasites and host cells. Noteworthy, intracellular calcium ion levels are essential for secretion and function of T. gondii MICs in parasites [273841].
The MICs are recognized by specific receptors on the cell membrane of hosts. Various methods (such as the proteomic and genomic approaches) have been used to detect the contents of the micronems in apicomplexan parasites. The electronic-microscope was shown micronemes are composed of electron-dense matrix due to the high protein content with secretory organelles that are important for gliding [3842]. Overall, depending on the species of parasite and the developmental stages, the MIC family includes at least 19 types in mammals (MIC1-12, M2AP, AMA1, ROM1, PLP1, SUB1, TLN4, and SPATR), of which 10 types have been identified with adhesive motifs such as epidermal grow factor (EGF) and chitin binding-like. Noteworthy, these products are essential for adhesion to host cells by parasites [4344]. However, MICs became famous as effective vaccine candidates against T. gondii, due to their basic roles in the early stage of the invasion of host cells by parasites. In this field, many kinds of MICs such as MIC2, MIC3, MIC4, MIC8, MIC11, and MIC13 have been evaluated [40454647484950].
It has been recognized that MIC11 as a soluble protein containing a α-chain and a β-chain tethered by a disulfide bond that take part the early stage of cell invasion. These data suggested that MIC11 protein is able to provoke the humoral and Th1-type immune responses, significant enhancement of interferon-γ (IFN-γ), interleukin (IL)-12, and IL-2 production along with higher survival time, compared to control groups [50], suggesting it could be a potential vaccine candidate. MIC8 is a promising vaccine candidate against acute and chronic toxoplasmosis infection that is expressed in tachyzoite stage of the life cycle and acts as an escorter for soluble adhesions to the cells. Notably, this protein plays a crucial role during the invasion of the parasite into the host cell as well as involved in the intracellular proliferation of parasite. Also, it is introduced as a potent stimulator for specific immune responses [4851]. An excellent article indicated favorable and promising results for MIC8 including increased humoral and cellular immune responses with the predominance of IgG2a over IgG1 (T helper 1 [Th1]-polarized responses), enhanced number of CD4+ and CD8+ T cells (p<0.05), high production of IFN-γ, prolonged survival time, and significant reduction in percentage of brain cyst load [51]. The specific features and main functions of some MICs have been listed in Table 1.
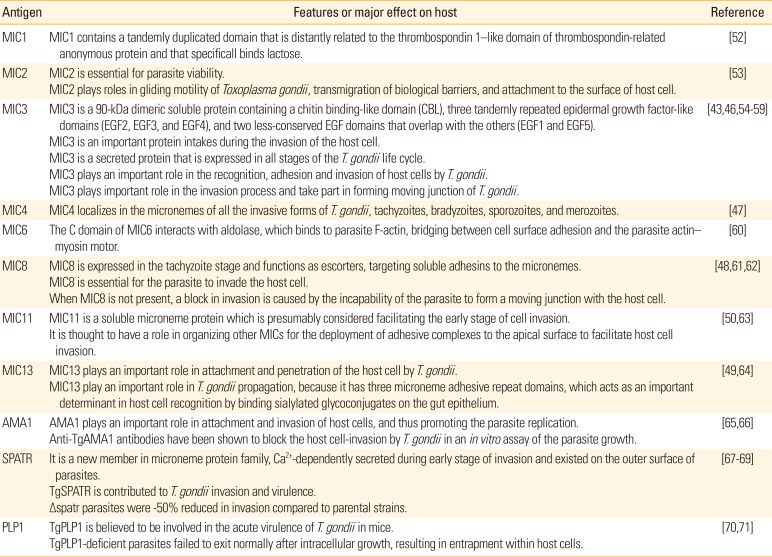
Table 1
The main features and functions of some MICs
| Antigen | Features or major effect on host | Reference |
|---|---|---|
| MIC1 | MIC1 contains a tandemly duplicated domain that is distantly related to the thrombospondin 1-like domain of thrombospondin-related anonymous protein and that specificall binds lactose. | [52] |
| MIC2 | MIC2 is essential for parasite viability. | [53] |
| MIC2 plays roles in gliding motility of Toxoplasma gondii, transmigration of biological barriers, and attachment to the surface of host cell. | ||
| MIC3 | MIC3 is a 90-kDa dimeric soluble protein containing a chitin binding-like domain (CBL), three tandemly repeated epidermal growth factor-like domains (EGF2, EGF3, and EGF4), and two less-conserved EGF domains that overlap with the others (EGF1 and EGF5). | [4346545556575859] |
| MIC3 is an important protein intakes during the invasion of the host cell. | ||
| MIC3 is a secreted protein that is expressed in all stages of the T. gondii life cycle. | ||
| MIC3 plays an important role in the recognition, adhesion and invasion of host cells by T. gondii. | ||
| MIC3 plays important role in the invasion process and take part in forming moving junction of T. gondii. | ||
| MIC4 | MIC4 localizes in the micronemes of all the invasive forms of T. gondii, tachyzoites, bradyzoites, sporozoites, and merozoites. | [47] |
| MIC6 | T he C domain of MIC6 interacts with aldolase, which binds to parasite F-actin, bridging between cell surface adhesion and the parasite actin–myosin motor. | [60] |
| MIC8 | MIC8 is expressed in the tachyzoite stage and functions as escorters, targeting soluble adhesins to the micronemes. | [486162] |
| MIC8 is essential for the parasite to invade the host cell. | ||
| When MIC8 is not present, a block in invasion is caused by the incapability of the parasite to form a moving junction with the host cell. | ||
| MIC11 | MIC11 is a soluble microneme protein which is presumably considered facilitating the early stage of cell invasion. | [5063] |
| It is thought to have a role in organizing other MICs for the deployment of adhesive complexes to the apical surface to facilitate host cell invasion. | ||
| MIC13 | MIC13 plays an important role in attachment and penetration of the host cell by T. gondii. | [4964] |
| MIC13 play an important role in T. gondii propagation, because it has three microneme adhesive repeat domains, which acts as an important determinant in host cell recognition by binding sialylated glycoconjugates on the gut epithelium. | ||
| AMA1 | AMA1 plays an important role in attachment and invasion of host cells, and thus promoting the parasite replication. | [6566] |
| Anti-TgAMA1 antibodies have been shown to block the host cell-invasion by T. gondii in an in vitro assay of the parasite growth. | ||
| SPATR | It is a new member in microneme protein family, Ca2+-dependently secreted during early stage of invasion and existed on the outer surface of parasites. | [676869] |
| TgSPATR is contributed to T. gondii invasion and virulence. | ||
| Δspatr parasites were -50% reduced in invasion compared to parental strains. | ||
| PLP1 | TgPLP1 is believed to be involved in the acute virulence of T. gondii in mice. | [7071] |
| TgPLP1-deficient parasites failed to exit normally after intracellular growth, resulting in entrapment within host cells. |
![]()
DNA Vaccines
During two recent decades, continuous efforts of researchers have made precious achievements in the development of DNA vaccines against acute and chronic toxoplasmosis [2429]. DNA vaccine as a robust strategy has been developed instead of traditional approaches, because of the following reasons: long-term persistence of immunogenicity, relative stability, absence of any type of microorganism, high safety, cost benefits, ease of handling, etc. [277273].
DNA vaccines in general have shown to be effective for inducing both humoral and cell mediated immune response and also stimulates dendritic cells (DCs) to be matured and makes them strong stimulators of T-cell immunity [242772]. Interestingly, activation of B-cells prevent from the attachment of T. gondii to its host cell receptors depend on the production of specific antibodies that can eliminate the parasite with the help of macrophages (MQs) [74]. As both humoral and cellular immunity responses stimulate during toxoplasmosis infection, Th1 immune response has a critical role to limit the parasite replication and produce cytokines such as IL-2 and IL-12. Also, IL-12 is produced by innate immune cells such as DCs, MQs, neutrophils, and monocytes, which plays an important role in host resistance [24757677]. The natural killer (NK) cells and T-CD8+ and T-CD4+ seem to be an important source of IFN-γ in the early and chronic phases of infection. This cytokine as the adaptive cellular immunity has a key role in the controlling and restriction of the parasite as well as inhibit the reactivation of bradyzoites inside the dormant tissue cysts [7677]. In addition, Th2 cells produce IL-4, IL-5, and IL-10, which contribute to the regulation of cell-mediated immunity response reduction [78].
With a unique design strategy of DNA vaccine, it can induce cytotoxic T lymphocytes and helper immune responses with the cooperation of major histocompatibility complex pathways. Moreover, the immunity against T. gondii can be stimulated, so that several antigens can be detected from different epitopes at the same time [7273]. DNA vaccines can be injected through different routes such as, intramuscular, subcutaneous, mucosal, or transdermal [2773]. After the injection, the naked DNA plasmid enter to the cell cytoplasm in order to express encoded proteins within the host cells. As a result, induces a strong immune response [72].
Recent studies showed that the use of genetic and non-genetic adjuvants have become popular in order to provide sufficient immunity [24272979]. It should be mentioned that adjuvants have an important role to improve the efficacy of a vaccine by enhance either the magnitude or time of DNA expression and recruiting the immune cells to the site of injection. Also, they are used in order to help the uptake of DNA into host cells as well as increase taken up by professional antigen-presenting cells [2772]. Several publications have demonstrated that cytokines such as (IFN-γ, IL-12, IL-15, IL-21, etc.), chemokines, and costimulatory molecules (B7-1, B7-2, etc.) as adjuvant, could boost the effectiveness of DNA vaccines [2433465171798081]. For instance, IL-21 synergizes with IL-15 to enhance the generation of CD8+ memory T cells and NK cell activity. Thus, these cytokines can suggest as a candidate adjuvant against toxoplasmosis [51]. For this purpose, Li et al. (2014) [51] designed an investigation to evaluate the immunogenicity of pVAX-MIC8 plus pVAX/IL-21/IL-15. The findings showed co-administration of MIC8 plus mIL-15 and m-IL-21 cytokines enhanced survival time and improved protective immunity of DNA vaccine [51]. Noteworthy, as a member of germ line-encoded receptor, toll-like receptors (TLRs) have a special ability in the innate and adaptive immune responses to pathogens. Hence, they are the target of new vaccine adjuvants in order to improve the immunogenicity of DNA vaccines [82]. For instance, oligodeoxynucleotides contained CG motifs (CpG ODN) as the TLR-9 ligand and a molecular adjuvant to be effective to enhance the immunogenicity of DNA vaccines [7283].
Recently, several papers have examined the various MICs based on DNA vaccination approach, including MIC2, MIC3, MIC4, MIC6, MIC8, MIC11, MIC13, PLP1, M2AP, AMA1, and SPATR in different mouse models [3245464748495051545558596871848586878889]. Yuan et al. (2013) [49] evaluated the immunoefficacy of TgMIC13. The Kunming mice immunized with pVAX-TgMIC13 showed higher levels of IgG antibodies (p<0.05), T-cells proliferative response, high secretion of IFN-γ, IL-2, IL-4, and IL-10 (p<0.05), increased survival time (p<0.05), and significant reduction in the percentage of brain cysts load (p<0.05), compared with those mice that received phosphate-buffered saline. These data suggest that T. gondii MIC13 is a reasonable vaccine candidate against acute and chronic T. gondii infection [49]. MIC3 as a secreted protein plays an essential role in the attachment and invasion of host cells, which expressed at all stages of the life cycle of T. gondii (tachyzoite, bradyzoite, and sporozoite) and discharged from small secretory vesicles [84]. It has been shown that CBA/J mice vaccinated with a plasmid encoding of MIC3 (pMIC3i) produced a significant cellular immune response with the increased secretion of IFN-γ and IL-2 cytokines. The findings showed the response was increased by the pMIC3i plus the plasmid encoding granulocyte-macrophage colony-stimulating factor (pGM-CSF). Also, the immunized mice showed a dramatic reduction of brain cyst load against an oral challenge with T. gondii 76K cysts, compared with control mice [46]. It has been reported that CD4+ and CD8+ T lymphocytes have a key role in MIC3 DNA vaccine to induce protection. Furthermore, plasmids encoding the EGF-like domains and the Lectin-like domain of MIC3 are involved in the protection [90]. More examples of immunization experiments with DNA vaccines against T. gondii in different mouse models are listed in Supplementary Tables 1 and 2.
Recently, it has been well established that using a combination of multiple antigens to be more effective compared with single antigens, as well as improve the protective immunity against toxoplasmosis either survival duration time and/or brain cyst load [325458]. For instance, Beghetto et al. (2005) [89] evaluated five distinct protein fragments MIC2, MIC4, M2AP, and AMA1 gene products that are recognized by antibodies and T cells from infected individuals. The highest protection with DNA vaccination against T. gondii infection was obtained by immunization of BALB/c mice with plasmid mixture and the brain cyst burden in mice vaccinated with the various proteins was significantly reduced than those in control groups. The authors concluded that microneme gene fragments with the antigenic regions of cyst-specific genes could be useful in vaccination against toxoplasmosis [89]. In another study, Fang et al. [85] examined MIC3 and SAG1 proteins alone or combined together. The BALB/c mice were intraperitoneally immunized with 1×103 tachyzoites of RH strain. The results revealed that single-gene immunization increased humoral immune responses, increased secretion of IFN-γ, and prolonged the survival time, compared with the control groups (p<0.05). On the other hand, those mice that vaccinated with the multi-antigenic DNA vaccine (MIC3/SAG1), boosted the protective immunity in terms of cytokine production and survival time, compared with single gene immunized groups (p<0.05). These observations led to the suggestion that MIC3 with SAG1 together capable to induce long term and significant protection against toxoplasmosis, and also cocktail-vaccine immunization could be employed as an alternative way to providing effective protection against T. gondii infection [85].
Recombinant Protein Vaccines
Over the past decades, considerable progress has been achieved to recognize the molecular biology of the various aspects of T. gondii that resulted to design of different vaccine experiments against toxoplasmosis, based on the subcellular components of the parasite [273843919293]. The family of micronemes are attractive vaccine candidates that are responsible for the host-cell invasion [4353]. One of the alternative ways for the development of vaccine candidates against toxoplasmosis is recombinant subunit vaccines that have high potency to trigger systemic humoral and cell mediated responses as well as they are very important for large-scale production [4094].
MICs promote Th1 response, which is critical in mediating the resistance to T. gondii. In order to evaluate the protective efficacy of recombinant form of MICs, Pinzan et al. (2015) [40] designed a comprehensive study on different recombinant microneme proteins (TgMIC1, TgMIC4, and TgMIC6) and combinations of these proteins (TgMIC1-4 and TgMIC1-4-6). They vaccinated the C57BL/6 (H-2b) mice subcutaneously with TgMIC1 (10 µg), TgMIC4 (10 µg), TgMIC6 (10 µg), TgMIC1-4 (5 µg of each protein), TgMIC1-4-6 (3.3 µg of each protein), or Lac+ (10 µg) emulsified in Freund's complete adjuvant. One month after the last immunization procedure, the mice were orally infected with 40 and 80 cysts of the ME49 strain for chronic and acute toxoplasmosis, respectively. The results indicated that these recombinant protein vaccines significantly enhanced IgG titers, mixed Th1/Th2 responses with the predominance of IgG2b over IgG1, high production of IFN-γ and IL-10 cytokines with strong lymphocyte proliferative responses, as well as the increased survival rate (p<0.05), compared with control groups. Besides, immunization with TgMIC1-4 and TgMIC1-4-6 vaccines boosted the protective efficiency, so that 70% and 80% of immunized mice survived 30-day post challenge, respectively. The brain cyst load in mice vaccinated with the different proteins was reduced than non-vaccinated groups ranging from 27.2%–67.8%. It is well known that multicomponent vaccine has better effects than single antigens. The authors declared that the use of this vaccine offers a promising strategy for conferring protection against toxoplasmosis [40]. More details can be found in Supplementary Tables 3 and 4.
Vaccines Based on Live-Attenuated Vectors
Live-attenuated vectors such as bacteria or viruses are another strategy for enhancing the antigen presentation to the immune system of the host. They can greatly mimic the intracellular niche of T. gondii as well as provokes a strong humoral and cell mediated immune response, due to their intrinsic adjuvant properties [8595]. Also, these vaccines can be delivered by several routes, including intramuscular, intraoral, intranasal, subcutaneous, and intravenous in order to induce effective protection [96].
Recently, recombinant viral vectors have shown great potential and play a critical role to induce humoral and cellular immune responses. Hence, they could be a suitable vector for the development of new vaccines [859596979899]. For instance, it is well known that pseudorabies virus (PRV) has a high capability and remarkable effectiveness to enhance vaccine potency. The study showed that a new recombinant modified PRV expressing TgSAG1 (rPRV-SAG1) and TgMIC3 (rPRV-MIC3) cocktail induced a strong IgG antibody response and significant levels of IFN-γ, IL-2, and IL-10 production as well as increased the survival rate (66.7% survival 28 days, p<0.05) post challenge with 100 tachyzoites of RH strain in BALB/c mice. The authors remarked that expression of protective antigens of T. gondii in PRV is a novel approach towards the development of a vaccine against toxoplasmosis [97].
Virus like particle (VLP) vaccines are genetically engineered complexes of multiple copies of protein antigens in a particulate virus like structure that lacks viral genetic material and therefore cannot replicate [100]. These vaccine types have several advantages as follows [101]:
-Well-defined geometry and remarkable uniformity with repetitive and ordered surface structures
-Particulate and multivalent nature
-Preservation of native antigenic conformation
-Safety, as they are absolutely non-infectious and nonreplicating candidates
-Higher stability than soluble antigens in extreme environmental conditions
-Applicability as vectors for the presentation of foreign antigens
VLP vaccines can stimulate powerful humoral and cellular immune responses, representing one of the most appealing approaches for a vaccine platform by mimicking the main structural and functional characteristics of viruses [101102]. Moreover, they can be produced in insect cell expression systems, where foreign antigens can be displayed [102]. It has been reported that VLPs are being useful and safe as vaccine candidates that could provide stronger and longer-lasting protection against toxoplasmosis [98]. In this case, Lee et al. (2017) [98] reported a novel recombinant VLPs carrying MIC8 and then evaluated the immune response and survival status in BALB/c mice. Interestingly, MIC8 in VLPs able to elicit significantly both humoral and cell mediated responses. After immunization, the levels of IgG antibody in sera and IgA antibody in feces elevated by MIC8 VLP vaccine, compared than controls. The enhanced survival duration (intranasal group 100% protection and intramuscular group 60% protection 16 days after challenging with 1×105 tachyzoites of RH strain) was observed, compared with control mice that died within 12 days. Moreover, the numbers of germinal center B cell (B220+, GL7+) and T cell (CD4+, CD8+) populations increased more obviously, in the group immunized with MIC8 VLP than in the control group. These results provide an effective approach for developing vaccines based on VLPs for protection against the highly virulent RH strain of T. gondii [98]. Supplementary Table 5 listed the examples of immunization with live-attenuated vectors expressing T. gondii antigens in mouse models.
Prime-Boost Strategies
Over the past few years, prime-boost strategy has been evaluated for vaccine development, such as DNA prime/viral vector boost, DNA prime/protein boost, and protein prime/DNA boost that are able to induce both humoral and cell-mediated immune responses against many different pathogens especially intracellular pathogens [597299103104]. Hence, this strategy can be useful to promote the effectiveness of vaccine experiments.
Homologous prime-boost approach involves the similar formulation employed in both the prime and boost regimens, while heterologous prime-boost strategies contains different formulations used in more than one immunization [72105]. The interval between prime and boost is very substantial for vaccine response and high efficacy. Moreover, the arrangement of vaccination schedule undoubtedly could influence the outcome of prime-boost strategies [72106]. Noteworthy, heterologous prime-boost is more likely to be immunogenic against an antigen than the homologous prime-boost [105107]. The advantage of heterologous prime-boost immunization is the induction of a strong cellular immune response and is associated with a higher and more specific antibody response against the vaccine target compared to homologous immunization. It has been reported that in comparison with homologous prime-boost approach with the same DNA vaccine, boosting a primary response with a heterologous vector leads to 4- to 10-fold higher T-cell responses [105]. A DNA or a viral vector, especially adenovirus for priming and a protein-based vaccine as a booster, have been used in heterologous prime-boost strategy [107]. Several different vaccine strategies have been shown to elicit different types of immune response. For example, DNA vaccines or recombinant live vector-based vaccines are able to elicit an efficient cellular immunity, but subunit vaccines commonly trigger a predominant humoral immunity [99105]. Subunit vaccines are based on peptides, proteins or polysaccharides containing protective antigens. However, the recombinant subunit vaccines are poorly immunogenic and usually require some additional components to enhance the potency of protective immunity. Accordingly, the use of some adjuvants and also repeated boost immunizations are suggested to elevate the efficiency of subunit vaccines [105108]. The underlying mechanisms of the effectiveness of prime-boost regimens still remain poorly understood. It is hypothesized that the lower antigen expression from DNA vaccines may preferentially prime Th cell responses with the humoral response being subsequently boosted by the protein or viral vector boost [72]. The examples of heterologous prime-boost vaccination against T. gondii in mouse models have been inserted in Supplementary Table 6.
Conclusion
During the twenty years ago, the different vaccine types with various strategies have been evaluated throughout the globe. Despite of the continuous efforts of scientists, there is no available licensed vaccine for use in human and animals. Thus, the development of an efficient vaccine urgently required to prevent and limit the infection. The use of DNA vaccines encoding MIC2, MIC3, MIC11, M2AP, and AMA1 alone or as mixture increase the survival time/rate in immunized mice (see Supplementary Tables 1 and 2). Furthermore, the use of recombinant protein vaccines showed an elevated survival rate (up to 80% protection). Nevertheless, these investigations failed to report complete protection. The use of live-attenuated vectors as vehicles to deliver and express the target gene and prime-boost regimens are excellent strategies for vaccine development, which need more attention in the future studies. Reportedly, it has been verified that the use of genetic and non-genetic adjuvants is very effective, because of their potential ability in boosting specific and long-lasting protective immunity. Collectively, the obtained findings are widely diverse, but valuable advacements have been achieved, so that they gave promising perspectives for the future investigations. However, several limitations may affect the outcome of the experimental studies such as: unsuitable immunization protocol, inadequate evaluation criterion, the strain of parasite, evaluation criterion, the vaccine construct, dosage of inoculum, the delivery route, the different mouse models, etc. The future experiments must address the all these aspects to minimize the faults. In the other hands, optimize immunization protocol and use of various types of delivery systems, traditional and/or molecular adjuvants undoubtedly would affect the outcomes.
 XML Download
XML Download