

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Cholera is a global public health problem, especially in developing countries. Global burden of cholera was estimated 1.4 to 4.0 million cases and 21,000 to 143,000 individuals died annually in the world [1]. Since the disease is generally transmitted by contaminated water and food, access to safe drinking water and sanitation in vulnerable communities is fundamental approach to preventing the disease [2]. Currently, vaccination is the most efficient way to reduce cholera infection in endemic area as short- to mid-term strategy [3]. Three oral cholera vaccines (OCVs), Dukoral, Shanchol, and Euvichol, have been prequalified by the World Health Organization (WHO) for use in global public health. Among them, Shanchol and Euvichol were manufactured according to the same formulation developed and technology transferred by the International Vaccine Institute (IVI), which contains heat or formalin killed Vibrio cholerae O1 Inaba, O1 Ogawa, and O139 with thimerosal (TM) as preservative.
TM, mercury-based preservative, has been widely used in medicinal drugs and vaccines [4]. There is no direct evidence on adverse effect of low doses of TM in vaccines. However, based on data regarding the toxicity of a related substance, methylmercury, the theoretical potential for neurotoxicity caused by low level of organomercurial compounds has been addressed as a concern [56]. Therefore, vaccine manufactures and regulatory agencies in Europe and North America recommended to reduce or eliminate TM in the vaccines for safety in 1999 [7]. The WHO issued guidelines on regulatory expectations related to the elimination, reduction or replacement of TM in vaccines [8] while it continues to recommend the use of vaccines containing TM for global immunization programs, because it determined that the benefits of using such products far outweigh any theoretical risk of toxicity [9].
After the OCV Shanchol, containing TM, received WHO prequalification in 2011, due to the increasing OCV demand and unmet need in the public market, a second technology transfer from IVI to a Korea based company, EuBiologics Co. Ltd., led to the manufacturing of Euvichol with the same formulation as Shanchol. Euvichol received WHO prequalification in 2015. Since then, the OCV stockpile for use by public health agencies like WHO and United Nations Children's Fund (UNICEF) was dramatically expanded. Here, we investigated in an animal model whether TM-free Euvichol could induce protective immunity and antibody responses against Vibrio cholerae comparable to the TM-containing formulation.
Go to : 
Materials and Methods
Bacterial strain and reagents
V. cholerae O1 El Tor Inaba strain, T19479 was used for vibriocidal assay and bacterial challenge study. TM-containing Euvichol (Lot. No. 14001) and TM-free Euvichol (Lot. No. TF-15002) were kindly provided by EuBiologics Co. Ltd., Chuncheon, Korea
Immunization
All experiments were performed with approval of institutional animal care and use committees of the International Vaccine Institute (IACUC approval No. 2016-003). Seven-week-old female BALB/c mice were purchased from Koatech (Pyeongtaek, Korea). All mice were acclimated for 1 week before use. Immunization and bacterial challenge studies were carried out as previously described [10]. Briefly, mice (n=10-12 per group) were stratified into five groups: non-immunized, immunized via oral or intranasal route with either of the vaccines (TM-containing or -free Euvichol). Mice were administered with 150 µL, equivalent to 10% of vaccine dose, of Euvhichol via oral route or 15 µL, equivalent to 1% of vaccine dose, via intranasal route on days 0 and 14. For bacterial challenge, mice were infected intranasally with 2.5×107 colony-forming unit (CFU) of V. cholerae strain T19479 at 1 week after the last immunization. Blood samples were collected from non-immunized and immunized mice groups on day 21 and serum samples were separated following blood clotting at room temperature (RT) for 2 hours.
Enzyme-linked immunosorbent assay
Anti-V. cholerae IgG, IgM, and IgA against V. cholerae in serum were measured by enzyme-linked immunosorbent assay as previously described [11]. Briefly, 96-well plate (Nunc, Cat.469454) were coated with 100 µL of diluted Euvichol (1:667) in phosphate buffered saline (PBS) at 4℃ overnight. After blocking with 300 µL of 1% bovine serum albumin (BSA) in PBS, serum samples were serially diluted with PBS containing 1% BSA from an initial 1:50 dilution and incubated for 1 hour at RT. The plates were washed with PBS containing 0.05% Tween 20 and incubated with horseradish peroxidase-conjugated anti-mouse IgG (γ chain specific, 1:5,000), anti-mouse IgM (µ chain specific, 1:5,000), or anti-mouse IgA (α chain specific, 1:5,000) at RT for 1 hour. After washing the plates, the wells were incubated with 100 µL of TMB substrate reagents for 20 minutes and 50 µL of 2 N H2SO4 to stop the reactions. Optical density was read at 450 nm using microplate reader. Endpoint titers were expressed as the reciprocal log2 of the highest dilution that gave an absorbance value of >0.1. The antibody titers below 50 were regarded as 25 for statistical analysis.
Vibriocidal assay
Vibriocidal assay was performed as previously described [12]. Briefly, complements in mouse serum was inactivated by heating at 56℃ for 30 minutes before use. Serum sample were 2-fold serially diluted with 0.85% saline from an initial 1:8 dilution and 25 µL of diluted serum was added into 96-well plate (Cat.269620, Nunc, Roskilde, Denmark). V. cholerae was cultivated in brain heart infusion (BHI) media at 37℃ for 1.5–2 hours until reaching mid-log phase. The cultured bacteria was washed and resuspended in saline containing 1×106 CFU/mL and 5% guinea pig complements (Rockland, Gilbertsville, PA, USA). Then, 25 µL of mixture of bacteria and complements was added to 96-well plate containing diluted serum. After incubation at 37℃ for 1 hour, 150 µL of BHI media per well was added and incubated at 37℃ for additional 4 hours. Optical density was read at 600 nm to measure bacterial growth using microplate reader. The vibriocidal titer was defined as the reciprocal log2 of highest dilution that completely inhibited bacterial growth. The vibriocidal antibody titers below 8 were regarded as 4 for statistical analysis.
Statistics
For comparison of vaccine-induced antibody responses, one-way ANOVA was carried out, followed by Tukey's multiple comparison, with significance determined at p<0.05 using GraphPad Prism 5 software (GraphPad Software, La Jolla, CA, USA).
Go to :
Results
Intranasal immunization with TM-free and -containing Euvichol protects mice against V. cholerae challenge
To examine whether immunization with TM-free and -containing Euvichol could protect mice against V. cholerae challenge, mice were immunized with either of the two Euvichol by oral or intranasal route on days 0 and 14. One week after the last immunization, mice were intranasally challenged with a lethal dose (2.5×107 CFU) of V. cholerae O1 Inaba. As shown in Fig. 1A, all non-immunized mice succumbed within 24 hours after challenge. Oral immunization with Euvichol, regardless of the presence of TM, could not provide protection to mice against V. cholerae challenge. However, all mice intranasally immunized with TM-containing and -free Euvichol were protected against bacterial challenge until 10 days post infection. Approximately, 5% of body weight was decreased at 1 day after V. cholerae challenge, but mice fully recovered their weight loss from the next day (Fig. 1B). These results suggest that TM-free Euvichol induce protective immunity against V. cholerae comparable to the TM-containing formulation in cholera pulmonary infection model.
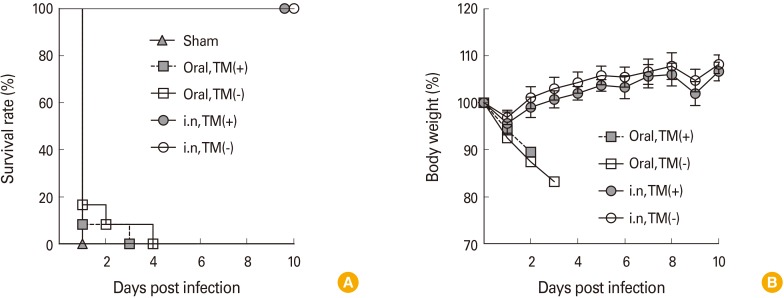 | Fig. 1Intranasal immunization with thimerosal (TM)-free and -containing Euvichol protects mice against a challenge with Vibrio cholerae O1 Inaba. Four groups of mice (n=10-12) were administered intranasally or orally with TM-containing or -free Euvichol on day 0 and day 14 against a control group of non-immunized mice. Survival rate (A) and percentage weight change (B) were daily monitored following challenge with a lethal dose of V. cholerae O1 Inaba. i.n., intranasal; TM(+), thimerosal-containing; TM(−), thimerosal free.
|
Intranasal immunization with Euvichol elicited strong systemic antibody responses against V. cholerae
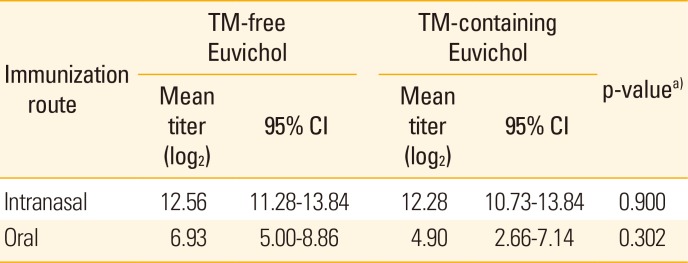
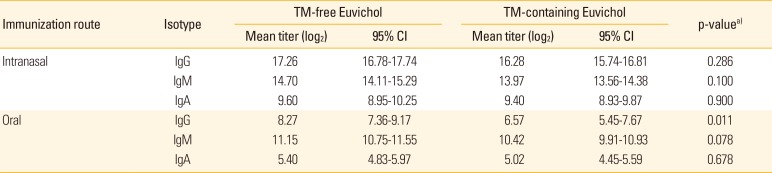
Serum vibriocidal antibody is widely used to evaluate immunogenicity of cholera vaccine in clinical trials as surrogate marker [13]. To assess the systemic antibody responses induced by immunization with Euvichol, we measured vibriocidal activity in serum obtained from mice administered with TM-containing or -free Euvichol (Table 1, Fig. 2). Sera from mice intranasally immunized with both variations of Euvichol showed significantly higher vibriocidal titers against V. cholerae O1 Inaba than those from orally immunized mice (TM-free, intranasal 12.56 vs. oral 6.93, p=0.001; TM-containing, intranasal 12.28 vs. oral 4.90, p=0.001). When analyzing separately the orally and the intranasally immunized groups, there was no significant difference in vibriocidal titers against V. cholerae O1 Inaba between TM-free and -containing Euvichol in both oral (12.56 vs. 12.28, p=0.900) and intranasal (6.93 vs. 4.90, p=0.302) immunization groups, respectively. In addition, antibody responses to V. cholerae were also assessed following immunization by oral or intranasal route (Table 2, Fig. 3). When mice were immunized with the TM-free formulation, vaccine-specific IgG, IgM, and IgA levels were significantly higher in intranasally immunized mice compared to orally immunized mice, (intranasal 17.26 vs. oral 8.27, p=0.001 for IgG; intranasal 14.70 vs. oral 11.15, p=0.001 for IgM; intranasal 9.60 vs. oral 5.40, p=0.001 for IgA). Similar trends in antibody responses between oral and intranasal route groups were observed when mice were immunized with TM-containing formulation. Interestingly, TM-free Euvichol generated statistically significant increase of serum IgG compared to TM-containing Euvichol in oral immunization group (8.27 vs. 6.57, p=0.011) but not in intranasal immunization group (17.26 vs. 16.28, p=0.286). There was no significant difference in the level of IgM and IgA between the two vaccine variations in either intranasal (14.70 vs. 13.97, p=0.100 for IgM; 9.60 vs. 9.40, p=0.900 for IgA) or oral immunization group (11.15 vs. 10.42, p=0.078 for IgM; 5.40 vs. 5.02, p=0.678 for IgA). Altogether, these data suggest that TM-free Euvichol could induce similar antibody responses to TM-containing Euvichol.
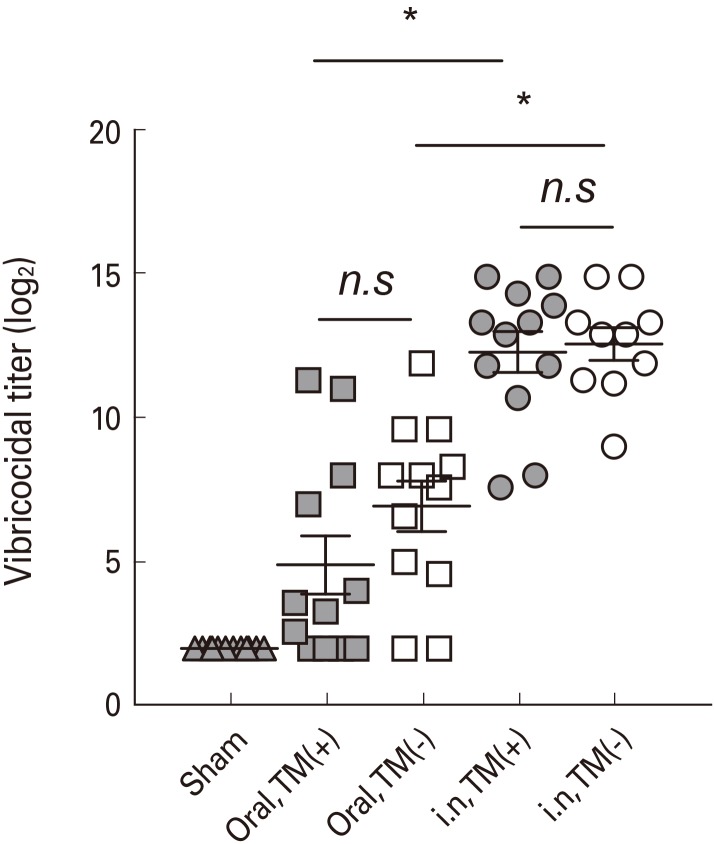 | Fig. 2Intranasal immunization with thimerosal-free and -containing Euvichol induces vibriocidal antibody response to Vibrio cholerae. Four groups of mice (n=10-12) were administered intranasally or orally with TM-containing or -free Euvichol on day 0 and day 14 against a control group of non-immunized mice. Serum samples were collected from non-immunized and immunized mice groups on day 21 and vibriocidal activities were measured. Data represent mean values±standard error of mean from triplicate assays. *p<0.05 indicates statistically significant difference between two experimental groups by use of one-way ANOVA with Tukey's multiple comparison test. n.s, not significant; i.n., intranasal; TM(+), thimerosal-containing; TM(−), thimerosal free.
|
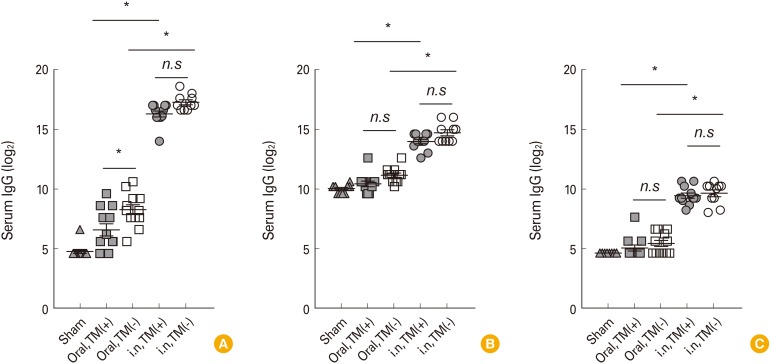 | Fig. 3Intranasal immunization with thimerosal-free and -containing Euvichol induces Vibrio cholerae-specific antibody responses. Four groups of mice (n=10-12) were administered intranasally or orally with TM-containing or -free Euvichol on day 0 and day 14 against a control group of non-immunized mice. Serum samples were obtained at 1 week after the last immunization, and V. cholerae-specific IgG (A), IgM (B), and IgA (C) titers were measured. Data represent mean values±standard error of mean from triplicate assays. *p<0.05 indicates statistically significant difference between two experimental groups by use of one-way ANOVA with Tukey's multiple comparison test. n.s, not significant; i.n., intranasal; TM(+), thimerosal-containing; TM(−), thimerosal free.
|
Table 1
Serum vibriocidal titers against Vibiro cholerae O1 Inaba
![]()
Table 2
Euvichol-specific serum IgG, IgM, IgA titers (log2)
![]()
Go to :
Discussion
TM has long been used in vaccines, especially in multi-dose formulation as preservative. Although there is no direct evidence of harmful effect caused by TM on human, public health sector and vaccine manufacturers are trying to remove it from products due to theoretical safety concerns based primarily on data regarding the toxicity of a related substance, methylmercury including potential risk for neurotoxicity [58]. In the present study, we evaluated the immunogenicity of TM-free OCV, Euvichol, in a murine pulmonary cholera model. Intranasal immunization with both TM-free and -containing Euvichol completely protected mice against V. cholerae challenge and induced strong systemic immune responses including vibriocidal antibody and vaccine-specific IgG, IgM, and IgA in serum. However, oral immunization could not provide protective immunity and antibody responses were less pronounced compared to intranasal immunization in mice model. These results are consistent with our previous study in mice intranasally administered with OCV Dukoral, which is comprised of whole-cell inactivated V. cholerae O1 and recombinant cholera toxin B subunit, where immunized mice were completely protected against cholera-induced pneumonia [10]. Given the fact that intranasal administration elicited significantly strong systemic antibody responses but not oral immunization, antibacterial antibodies may play a role in protection against V. cholerae infection. Local immunity may also contribute to protection, as observed in our previous study [10], although this was not observed in this study. In addition, the presence of TM in the vaccine did not affect cholera vaccine-induced immune responses in mice.
Serum vibriocidal antibody has long been considered as the best surrogate marker for protection against cholera [1415]. Intranasal administration, but not oral immunization, with OCVne evoked strong vibriocidal antibody response against V. cholerae O1 Inaba in the present study. This result is consistent with previous reports [1016]. Euvichol is composed of inactivated V. cholerae O1 Inaba, O1 Ogawa, and O139 and therefore, vibriocidal activities against all of three strains have been evaluated in clinical studies. However, in this animal study we based the comparison of immunogenicity between the two variations of Euvichol on O1 Inaba-specific vibriocidal antibody in consideration of the cross-reactivity between Inaba and Ogawa strain [17], and of the poor responses against O139 strain in previous studies [1819]. Vaccine-specific serum IgG, IgM, and IgA antibodies are considered to mediate protection against cholera infection in this model. This is supported from previous studies where LPS-specific serum IgG and IgA were highly increased in cholera patients and vaccinated individuals [202122]. In addition, serum IgM and secretory IgA against V. cholerae LPS generated in patients play a role in conferring protective immunity against cholera [2324].
Since V. cholerae does not induce natural infection in animals except for human, there is no accurate model to reflect the immune reaction occurred in clinical settings. Several animal models have been developed to evaluate cholera vaccine in pre-clinical stage. Infant mouse model has been widely used, but immaturity of sucking mice's immune systems and microflora does not allow to evaluate active immunity against bacterial challenge [25]. Germ-free mouse model could be used, because V. cholerae can colonize in the gut [26]. However, the cost for maintenance of germ-free animal is expensive and immunity is not fully developed due to lack of intestinal microbiota. In addition, antibiotic-resistant adult mice model has been developed, but the animal should be pre-treated with streptomycin to remove gut microflora, and streptomycin-resistant V. cholerae can only be used to examine bacterial colonization [16]. Overall, it is difficult to assess protection against natural infection with wild-type V. cholerae in this model. The mouse pulmonary cholera model used in this study seems appropriate for evaluation of immunogenicity of cholera vaccines in several aspects: use of adult mice is inexpensive and convenient to manipulate; well-characterized in immune system; protection against wild-type V. cholerae can be assessed. Nevertheless, there are limitations to evaluate the immunogenicity of cholera vaccines in correlation with the human. For instance, cholera infection in human is mainly by oral route and colonizes the small intestine, but, in the present study, only immunization by intranasal route showed successful protection against V. cholerae and fully induced antibody responses.
In summary, this study showed that the TM-free variation of the OCV Euvichol is highly immunogenic and induce similar immune responses compared to TM-containing Euvichol in mice. Following this study, in September 2016, the TM-free Euvichol obtained WHO prequalification. Subsequently, clinical data became available supporting the equivalence for safety and immunogenicity between the two Euvichol variations [27]. In conclusion, the pulmonary mouse cholera model represents a good candidate to examine in vivo the potency of cholera vaccines.
Go to :
 XML Download
XML Download