

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis (AD) is a chronic recurrent inflammatory skin disease characterized by itching and xerosis.1 The prevalence of AD is increasing in developed countries, with rates of 15% to 30% in children and 2% to 10% in adults.234 Epidemiological studies conducted in Korea have also indicated a similar prevalence and an increasing trend for AD.56 The pathogenesis of AD principally involves systemic and local factors. Systemically, AD is a cutaneous manifestation of a systemic disorder that is accompanied by other allergic disease, such as asthma and allergic rhinitis. Increased blood eosinophil, serum immunoglobulin E (IgE) and deviated systemic Th2 immune responses are characteristics of these systemic disorders.78 Locally, skin barrier dysfunction, immunologic triggers (allergen and microbial infection) and decreased skin innate immune response are included in the pathogenesis of AD.7 AD is likely to occur due to the interactions between systemic and local pathogenic factors.
The collective microbial community has been implicated in AD. Previous work showed that microorganisms affect systemic dysregulation of the innate and acquired immune systems. To the best of our knowledge, no study has comprehensively analyzed systemic microbiome factors. Identifying microorganisms in the blood or urine was considered to be futile, since these sites should be sterile; however, bacterial extracellular vesicles (EVs) have been observed in the blood and other organ systems and are known to contribute to the inflammation process.9 EVs are nanometer-sized vesicles derived from archaea, bacteria and eukaryotic cells that enclose cell components including proteins, nucleic acids and lipids, which have been implicated as having a physiological or pathological key role in cell communication.1011 Analysis of bacterial EV in the blood or urine can be exploited to study systemic bacterial conditions.12
The lack of studies concerning the overall microbiome has precluded a deep understanding of the effect of bacteria. To understand the complex relationship between human health and bacteria, an approach that is more sophisticated and comprehensive than the culture method is needed.13 Metagenomic analysis is not burdened by culture-related limitations because it analyzes bacterial DNA collected directly from the environment regardless of cell viability.1314
In the present study, we evaluated the systemic bacterial EV composition using metagenomic analysis of bacterial DNA in the urine of AD patients and normal controls. The goal was to identify bacteria contributing systemically to AD, to characterize changes in bacterial composition according to AD treatment and to elucidate the effect of EV from beneficial bacteria through in vitro and in vivo examinations.
MATERIALS AND METHODS
Study subjects
Patients visiting the Pediatric Allergy and Respiratory Center of Soonchunhyang University Hospital who had aggravated AD according to the Hanifin and Rajka AD diagnostic criteria15 were enrolled. Patients were excluded if they had other skin diseases, or had received topical, oral or injectable antibiotics within the previous 14 days. Wet dressing, topical corticosteroids and antibiotics were used to treat AD aggravation in all patients, and 66.7% and 18.5% of patients were treated with topical calcineurin inhibitors and systemic corticosteroids, respectively. Normal control children who were recruited had no skin diseases including AD and did not have any other allergic or significant underlying diseases. They were recruited from children who visited the Pediatric Department of Soonchunhyang University Hospital for vaccination and had not used probiotics and systemic antibiotics within 14 days. All participants were less than 17 years of age. Blood and urine were collected from the subjects. Participants and their parents provided informed consent. The study was approved by the Ethics Committee of Soonchunhyang University Hospital (approval number 2013-061).
Laboratory tests and measurement of AD severity
White blood cells and eosinophil count were measured in AD patients. The levels of total IgE, specific IgE and eosinophil cationic protein (ECP) were measured using ImmunoCAP testing according to the manufacturer's instructions (Phadia AB, Uppsala, Sweden). Laboratory tests were not performed in normal controls. AD severity was measured by the SCORing of Atopic Dermatitis (SCORAD) index16 by the same pediatric allergist to avoid inter-observer bias.
EV preparation from urine and serum, and DNA isolation
Multiple ultracentrifugation and purification processes were applied to isolate EVs, as described in a previous study.17 DNA was extracted using a commercial kit (Bioneer, Daejeon, Korea), and the quality and quantity of DNA were determined using spectrophotometry (NanoDrop Instruments, Wilmington, DE, USA).
Emulsion-based polymerase chain reaction (emPCR) and next-generation sequence analysis
After each clone was polymerase chain reaction (PCR)-amplified, the V1 to V3 regions were amplified using a FastStart High Fidelity PCR System (Roche Diagnostics GmbH, Mannheim, Germany) with a 16s rRNA gene fusion primer (27F-GAGTTTGATCMTGGCTCAG and 518R-WTTACCGCGGCTGCTGG). A GS-FLX plus emPCR Kit (454 Life Sciences, Branford, CT, USA) and Tissue Lyser II (Qiagen, Germantown, MD, USA) were used to make micro-reactors containing amplified compounds and beads. After emPCR amplification, the amplicon was purified using an AMpure Bead kit (Beckman Coulter, Brea, CA, USA) and quantified using the Picogreen method (Invitrogen, Carlsbad, CA, USA). Subsequently, the amplicon was diluted and analyzed with a GS-FLX Titanium sequencer (Roche Diagnostics GmbH) at Macrogen (Seoul, Korea).
Phylogenetic classification
Sequencing reads were obtained in high quality as determined by the Quality score (mean Phred score > 20 and read length > 300 bp). The operational taxonomic unit (OTU) was analyzed using UCLUST and USEARCH, with phylogenetic classification performed using QIIME based on the 16sRNA sequence database of GreenGenes 8.15.13. All sequences were classified based on the similarity as follows: species, > 97% similarity; genus, > 94% similarity; family, > 90% similarity; order, > 85% similarity; class, > 80% similarity; and phylum, > 75% similarity. If there was a significant difference (more than 2-fold difference between the case and control groups) in the bacterial EV composition of the genus level, then the data were configured as a heatmap.
Lactobacillus plantarum-derived EV isolation
We selected Lactobacillus in the mouse study because many studies have already confirmed the beneficial effects for eczema of Lactobacillus.
181920212223242526 Among Lactobacillus, the L. plantarum (KCTC 11401BP, Gen-Bank accession No. GQ336971) strain was isolated from kimchi, which is the most popular fermented food in Korea and is easily obtained.27 This strain also has the potential to improve mouse and human AD.2829 Finally, we decided to use L. plantarum for the experiment, and L. plantarum culture medium was donated by Probiotics Research Center, CJ CheilJedang Corporation, Korea. L. plantarum-derived EVs (Lp EV) were isolated as EV preparation methods.
Transmission electron microscopy (TEM) image analysis
Purified EVs were diluted to 50 µg/mL with phosphate-buffered saline (PBS), and 10 µL of the EV-suspension were placed on 300-mesh copper grids (EMS, Hatfield, PA, USA) and stained with 2% uranyl acetate on the 300-mesh copper grids. The microscope images were recorded with a JEM1011 electron microscope (JEOL Ltd., Tokyo, Japan).
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) analysis
The protein sample (10 µg) from EVs and whole cell lysate (WCL) were loaded into each well and analyzed with SDS-PAGE (10% resolving gel). The SDS-PAGE gel was stained with CBB R-250 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Dynamic light scattering
The purified EVs were diluted with PBS to 1 µg/mL and measured using a Zetasizer Nano ZS (Malvern Instruments, Worcestershire, UK) and Dynamic V6 software32.
In vitro evaluation of immunogenicity
Immortalized human epidermal keratinocytes (HaCaT cells) were donated by Jeung-Hoon Lee (Chungnam National University, Daejeon, Korea). The 2 × 105 HaCaT cells were seeded on 24-well plate in DMEM (Hyclone Laboratories Inc., South Logan, UT, USA) containing 5% fetal bovine serum (FBS) (Gibco, Grand Island, NY, USA) and 1% antibiotics (Hyclone Laboratories Inc.). Cells were treated with 0.1, 1 and 10 µg/mL EVs for 12 hours to evaluate the immunogenicity of Lp EVs. After treatment, IL-6 levels in the cultured media were evaluated via enzyme-linked immunosorbent assay (ELISA) following manufacturer's instructions (R&D Systems, Minneapolis, MN, USA).
In vitro protection of Lp EVs from Staphylococcus aureus-derived EVs
The cells were pretreated with Lp EV for 12 hours, followed by stimulation with S. aureus-induced EVs (Sa EV) for 12 hours. Co-treatment with Lp EV and Sa EV was performed to compare with the pretreatment group. After treatment, the cultured media was evaluated via ELISA by following manufacturer's instruction (R&D Systems). Some of the wells were treated for 24 hours and viability was measured using thiazolyl blue tetrazolium bromide (MTT; Sigma-Aldrich, St. Louis, MO, USA).
In vivo Lp EV administration on mouse skin inflammation model
Sa EVs were administrated to mouse skin 3 times per week for 4 weeks to develop the mouse model for skin inflammation. Before administration of Sa EV, the skin of each mouse was stripped 5 times using Durapore surgical tape (3M Co., St. Paul, MN, USA). Gauze (1 × 1 cm) was placed on the stripped mouse skin, and Sa EV in 100 µL PBS were sprayed onto the gauze, which was fixed to the skin using Tegaderm bio-occlusive tape (3M Co.). The Lp EV was administered orally 12 hours before each Sa EV administration for the Lp EV-treated group.
Measurement of cytokines in the mouse skin inflammation model
Skin tissue from each mouse was sampled to evaluate the production of IL-4 and IL-5 in the mouse model. Skin tissue samples were homogenized in RIPA Buffer (ThermoFisher Scientific, Waltham, MA, USA) and were centrifuged at 10,000 ×g for 20 minutes at 4°C. The supernatant was collected, and IL-4 and IL-5 levels were determined using ELISA according to the manufacturer's instructions (R&D Systems).
Histological analysis
Isolated skin tissues were fixed in paraffin, sectioned and stained with hematoxylin and eosin (H&E). The skin tissues were analyzed at 200× magnification.
Statistical analysis
The results of continuous variables are presented as the mean ± standard deviation. Continuous variables were analyzed using Student's t test or the Mann-Whitney U test, and categorical variables were analyzed using the chi-square test or Fisher's exact test. Statistical significance indicated by a P value of < 0.05. SPSS version 19.0 (SPSS Inc., Chicago, IL, USA) was used for analyses.
RESULTS
Clinical characteristics of AD patients and normal controls
Twenty-seven AD patients and 6 normal controls were enrolled. The mean ages of the AD patients and controls were 91 and 83 months, respectively. In both AD and control participants, 66.7% were boys. Eosinophil count and ECP were 877.8 cells/µL and 66.4 µg/L, respectively. Specific IgE to any allergen was positive in 26 patients. Among them, 66.7% were positive for the specific IgE for house dust mites. All patients were treated with wet dressings, topical corticosteroids, antihistamines and antibiotics. Topical calcineurin inhibitors and systemic corticosteroids were used in 66.7% and 18.5% of the patients, respectively. The SCORAD index was significantly decreased from 75.5 to 41.4 after treatment (P < 0.001). Laboratory tests were not conducted in the normal control group (Supplementary Table S1).
Comparison of bacterial EV composition in the urine and serum of the AD patients
Bacterial EV compositions in the urine and serum of the AD patients were compared each other to confirm the similarity of both sample types. The 2 sample types closely resembled each other in 18 AD patients whose serum and urine samples were collected simultaneously (Supplementary Fig. S1A and B). As shown in Supplementary Fig. S1C, the cluster of samples in blood (in blue) cannot be separated from the cluster of samples in urine (in orange), which implies that the bacterial compositions of the 2 groups are quite similar. This interpretation was confirmed further by the t test results with PC1 and PC2 (P value > 0.1). After confirming the similarity of bacterial EV composition in blood and urine by principal component analysis (PCA; Supplementary Fig. S1C), we decided to continue the analysis using urine samples, which were easier to collect.
Differences in bacterial EV composition in the urine of the AD patients and normal controls
Bacterial diversity as determined by the Chao 1 index was markedly decreased in AD patients compared to the control group (Fig. 1A). Definite separation of bacterial EV composition between AD patients and the control group was observed in PCA (Fig. 1B). At the phylum level, Firmicutes was dominant in the controls, and Proteobacteria was dominant in AD patients (Fig. 1C). A heatmap of the genus level revealed notable differences in bacterial EV composition in the urine of the control and AD patients (Figs. 1D and 2A). The proportions of Lactococcus, Leuconostoc, Lactobacillus and Lactobacillales(o) were significantly higher in the control group than in the AD patient group, and Alicyclobacillus, Propionibacterium and Streptophyta(o) were more increased in the AD patient group compared with the control group. Pseudomonas was common in both groups (Fig. 2A and B).
 | Fig. 1(A) Alpha-diversity as determined by the Chao 1 index. (B) Beta-diversity defined by PCA. (C) Bacterial community analysis of the urine of AD patients and normal controls at the phylum level. (D) Bacterial community analysis of the urine of patients and controls at the genus level (case: AD patients, control: healthy controls).PCA, principal component analysis; AD, atopic dermatitis.
|
 | Fig. 2(A) Differences in bacterial EVs composition in the urine of AD patients and normal controls. (B) Bacterial community analysis of the urine of AD patients and normal controls at the genus level.EV, extracellular vesicle; AD, atopic dermatitis.
|
Lactobacillus comprised 20.9% and 4.6% of the total flora in the control and AD patient groups, respectively. Respective values were 18.2% and 0.02% for Lactobacillales(o), 14.6% and 0.1% for Leuconostoc, and 11.6% and 0.03% for Lactococcus. Alicyclobacillus comprised 0.3% and 8.5% of the control and AD patient groups, respectively. Other respective values were 2.9% and 7.9% for Streptophyta(o), and 2.8% and 7.6% for Propionibacterium (Fig. 2B, Supplementary Table S2).
Post-treatment changes in the bacterial EV profile in urine in AD patients
Urine samples were collected before and after treatment from 10 AD patients. Then, we compared the changes in bacterial EVs composition based on the treatment in this group. Before the treatment, Alicyclobacillus and Comamonadaceae(f) were frequently found in AD patients. These proportions were decreased after treatment, while those of Acinetobacter and Oxalobacteraceae(f) were increased (Supplementary Fig. S2A and B). The proportions of Alicyclobacillus and Comamonadaceae(f) decreased from 8.7% to 1.9% and from 7.2% to 2.6%, respectively, after the treatment. The proportions of Acinetobacter and Oxalobacteraceae(f) were increased from 0.6% to 12.4% and from 3.0% to 9.2%, respectively, after the treatment (Supplementary Table S3). At the phylum level, Proteobacteria was dominant in both groups, and Firmicutes decreased after treatment. (Supplementary Fig. S2C).
Characterization of EVs from Lp EV
The Lp EV were purified according to EV isolation methods used in the previous study.26 First, we analyzed the morphology of EVs using TEM. We observed spherical lipid bi-layer vesicles (Fig. 3A). The size distribution of EVs was measured using a dynamic light-scattering analyzer. The diameter of EVs ranged from 20 to 100 nm (Fig. 3B). Next, we analyzed the protein patterns of EVs and WCL with SDS-PAGE. The EVs showed distinct protein patterns compared with WCL (Fig. 3C). These results suggest that EVs contain sorted proteins from host cells and that there are sorting mechanisms when EVs are produced.
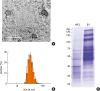 | Fig. 3Characterization of EVs from Lactobacillus plantarum. (A) TEM images of purified EVs from L. plantarum. (B) The size distribution of EVs from L. plantarum. The size distribution of EVs was measured in a diameter range of 20 to 50 nm. (C) SDS-PAGE analysis of purified proteins (WCL, EVs) from L. plantarum. Standard markers are shown on the left (kDa).EV, extracellular vesicle; TEM, transmission electron microscopy; SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis; WCL, whole cell lysate.
|
In vitro effects of Lp EV on the secretion of IL-6 from keratinocytes and macrophages
We assumed that EV from lactic acid bacteria would have some protective or beneficial roles against AD because we found that EVs of the lactobacilli group were dominant in the urine of normal controls compared to that of AD patients. As mentioned above, L. plantarum was selected as an experimental material among the lactic acid bacteria because it was easily obtained from kimchi, which is a familiar Korean food. First, we evaluated the therapeutic or preventive effect of EVs from L. plantarum on keratinocytes and macrophages. The concentration of the Lp EV used to treat the keratinocytes was determined at a level that did not cause skin irritation or inflammation compared to Sa EV, a positive control, which was already known to cause AD-like skin inflammation (Fig. 4A).3031 IL-6 secretion from keratinocyte was decreased with Lp EV treatment prior to Sa EV treatment in a dose-dependent manner, but not in the co-treatment model of Lp EV and Sa EV (Fig. 4A). IL-6 secretion from macrophages was also decreased with Lp EV pre-treatment, while no decrease was observed in the co-treatment group (Supplementary Fig. S3A). Cell viability was also restored with Lp EV pre-treatment on the keratinocytes (Fig. 4B). However, these trends failed to demonstrate statistical significance in macrophages (Supplementary Fig. S3B). These findings suggest that Lp EV could have a preventive effect on skin inflammation.
 | Fig. 4(A) In vitro effects of Lactobacillus plantarum on the secretion of IL-6 from keratinocytes stimulated by Sa EV. NC was treated only with PBS, and PC was treated only with Sa EV. Pre- and co-treatment indicates that Lp EV was treated before Sa EV stimulation, or at the same time, respectively. (B) Keratinocytes cell viability assessment.IL, interleukin; Sa EV, Staphylococcus aureus-induced extracellular vesicle; NC, negative control; PBS, phosphate-buffered saline; PC, positive control; Lp EV, Lactobacillus plantarum-derived extracellular vesicle.
*P < 0.05 vs. NC; †P < 0.05 vs. PC; ‡P < 0.01 vs. PC (Mann-Whitney U test).
|
In vivo effects of oral Lp EV on the AD mouse model induced by Sa EV
In the previous study, we demonstrated that Sa EV causes AD-like skin inflammation which was characterized by epidermal thickening with infiltration of the dermis by mast cells and eosinophils, and cutaneous production of IL-4, IL-5, interferon (IFN)-γ and IL-17.30 Epidermal thickening with the infiltration of inflammatory cells, especially eosinophils and increase in type 2 cytokines has been regarded as AD-like skin inflammation in mouse experiments.28303132 In the current study, we administered oral Lp EV to the Sa EV-induced AD mouse model to evaluate the in vivo effects of Lp EV on AD. Lp EV was administered orally for 25 days together with Sa EV application 3 times a week, and the effect was compared with that of dexamethasone treatment (Fig. 5A). Dexamethasone markedly decreased skin Sa EV-induced skin inflammation (Fig. 5B). Although low-dose Lp EVs (1 µg) were not effective in improving of skin inflammation, an increased dose reduced skin inflammation and showed an effect similar to the dexamethasone treatment (Fig. 5B). Histological analysis elucidated that eosinophilic inflammation and epidermal thickening were reduced after dexamethasone treatment. Lp EV administration also reduced epidermal thickening, but not eosinophilic infiltration (Fig. 5C). Elevated IL-4 levels in the positive control were significantly decreased with Lp EV administration (Fig. 5D). These results show that Lp EV may decrease skin inflammation by reducing IL-4 cytokine in a dose-dependent manner.
 | Fig. 5In vivo effects of oral Lp EV on the AD-like skin inflammation. (A) AD mouse model using tape-stripping and Sa EV. (B) Images of H&E-stained skin. (C) Histological analysis of epidermal thickness and infiltrating eosinophil. (D) Levels of IL-4 and IL-5.Lp EV, Lactobacillus plantarum-derived extracellular vesicle; AD, atopic dermatitis; Sa EV, Staphylococcus aureus-induced extracellular vesicle; H&E, hematoxylin and eosin; IL, interleukin; NC, negative control; PC, positive control.
*P < 0.05 vs. NC; †P < 0.05 vs. PC; ‡P < 0.01 vs. PC (Mann-Whitney U test).
|
DISCUSSION
The present study is the first comprehensive analysis of bacterial EV compositions undertaken to reveal the systemic proportion of bacterial EV in AD patients. Alicyclobacillus, Propionibacterium and Streptophyta(o) were increased in AD patients, and Lactococcus, Leuconostoc and Lactobacillus were significantly higher in the control group than in the AD patient group. These data indicate that a deficiency in systemic beneficial bacteria may promote development of AD. In AD patients, Alicyclobacillus and Comamonadaceae(f) were more frequent in urine before treatment, and Acinetobacter and Oxalobacteraceae(f) were more frequent after treatment. Then, we used L. plantarum to confirm the beneficial effects of lactic acid bacteria. In vitro examinations confirmed the anti-inflammatory effects of Lp EVs and their ability to prevent cell death. In vivo, Lp EV decreased epidermal thickening and IL-4 levels in the S. aureus EV-induced AD model. These findings suggest that Lp EV could have a preventive effect on skin inflammation.
Examinations of urine and blood samples involved detection of bacterial nano-vesicles, not the bacteria themselves. While bacteria might not be widely disseminated in the body, nano-vesicles derived from bacteria can be absorbed and move freely throughout the body.1233 Detection of bacteria-derived vesicles enabled analysis of systemic bacterial effects in humans for the first time. Recent data revealed microbiomes in organs or specimens that were considered sterile in the past. The microbiomes detected in the lungs or in urine are representative examples.910111234 Interestingly, there were notable differences in bacterial composition between urine in this study and skin in the previous study.35 Staphylococcus was found most commonly in the skin washing fluid of AD patients in our previous study.35 In contrast, Staphylococcus was not detected, and Alicyclobacillus, Streptophyta(o) and Propionibacterium were commonly found in the urine of AD patients. The bacterial community might be different between skin and systemic organ, even though Staphylococcus is the most common bacteria in the skin, their EV might not be absorbed systemically.
The EV of Propionibacteria, which was frequently observed in the urine of AD patients in this study, is not well known for its role in AD even though Propionibacteria is normal flora and is one of the most common skin colonizing bacteria.3637 Appropriate culture conditions are not yet established because this bacterium grows very slowly and is difficult to culture.38 Therefore, current culture methods are inappropriate to prove the relationship between Propionibacterium and AD. The present metagenomic results linking non-culturable bacteria with AD are important. Propionibacterium produces propionic acids and bacteriocin, which prevent proliferation of S. aureus or other pathogens. Therefore, it could play an opposing role relative to S. aureus.36 Therefore, Propionibacterium can be increased to counteract S. aureus in AD patients. Acinetobacter species are also known for their allergy-protective roles including those against asthma and AD.394041 This is consistent with our findings that an increased portion of Acinetobacter after the treatment of AD showed an inverse relationship with AD. In contrast, EVs of Pseudomonas may be commonly observed in the urine of both groups because they are part of the normal flora.42
There have been many studies about the beneficial effects of Lactococcus and Lactobacillus against allergic diseases.2122232425264344 Leuconostoc has not been studied frequently, but one study revealed that it can modulate IgE production in a mouse model with an ovalbumin-induced allergy.45 This result matches our observation that lactic acid bacteria were significantly higher in the urine of healthy controls than those of AD patients and showed protective effects both in vitro and in vivo. Leuconostoc has not been studied previously as a part of human microbiome in allergic diseases and our study is the first report of Leuconostoc in AD patients. Furthermore, we showed that not only bacteria itself, but also EVs from lactic acid bacteria might have the beneficial effects for AD.
However, the presence of other bacterial EVs in urine such as from Alicyclobacillus, Streptophyta(o), Comamonadaceae(f) and Oxalobacteraceae(f) needs to be studied further in order to determine their role and origin because there is little information about the EVs from these species. Our previous study revealed that Sa EV causes AD-like skin inflammation, and a successive study also showed that α-hemolysin in Sa EV plays an important role in inducing AD-like skin inflammation.4647 These studies strongly suggest a correlation between development of AD and bacteria-derived EV. Another study by the same research group showed that the degree of sensitization to the EV of dust is a risk factor for asthma, chronic obstructive pulmonary disease (COPD) and lung cancer.48 EVs might be involved in allergic inflammatory reactions of the lungs as well as skin. While there are relatively many studies on the relationship between allergic diseases and bacteria, few studies on EV derived from bacteria have been conducted. Therefore, further studies are needed in the future on allergic diseases and bacteria-derived EV.
There are some limitations in this study. First, the number of subjects is small, they are not representatives and the results cannot be applied to the general population. To generalize these results, large-scale studies and sample collection are essential. Secondly, it is still unclear whether lactic acid bacteria have any benefits to humans because we did not perform clinical trials. Their safety and efficacy should be evaluated to apply the treatment to human beings in the future. Thirdly, because AD patients in this study were treated with antibiotics when aggravated, the effects of antibiotics on the microbiome milieu should be considered when interpreting the differences after treatment.
Despite these limitations, our study is a breakthrough study in the bacterial EV composition analysis of AD patients. We analyzed the systemic bacterial composition through the study of nano-vesicles from bacteria and confirmed microbiome milieu by metagenomic analysis, which allowed a broader study of the whole bacterial community. Based on the culture technique to identify the bacteria, only 1% of the entire bacteria could be identified.49 Because metagenomic analysis provides much more information compared with a conventional culture technique, it is an essential method in the understanding of bacteria and human diseases.
In this study, we suggested the role of probiotics in the protection against AD. We also established the existence of novel bacterial EV in the urine reflecting a systemic microbiome milieu. Additional studies of microbiome might enable us to differentiate, diagnose and predict the prognosis of AD using microbiome information by local and systemic bacterial composition analyses. Furthermore, we might be able to prevent or treat AD using EVs from probiotics in the near future.
 XML Download
XML Download