

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Cancer is one of the leading causes of death worldwide and its incidence is dramatically increasing. The last few decades have witnessed significant progress in understanding of molecular and cellular basis of cancer cells. Metabolic alteration is one of the cancer hallmarks [1]. Metabolism in cancer cells is unique to support rapid proliferation, often making the environment under lack of nutrients and oxygen [234]. The cancer-specific metabolism provides a potential as therapeutic targets [5]. Serine metabolism is recently highlighted because of the roles in supporting tumor growth. Although the molecular mechanism is not fully understood, one possibility is through affecting production of S-adenosyl methionine (SAM), which is the substrate of DNA and histone methyltransferases. It suggests that serine metabolism directly or indirectly influence epigenetic alterations in cells [6]. Epigenetic mechanism contributes to altered gene function and cancer progression and its reversible nature has highlighted the application of epigenetic therapy [7]. Therefore, in this review, we focus on serine metabolism and its regulation by or on epigenetic modifiers in cancer cells.
CANCER METABOLISM
Cancer cells proliferate in 3 dimensional multicellular masses and the growing tumors as aggregation of cancer cells face insufficient supply of oxygen and nutrients. The hypoxic condition stimulates to form new blood vessels, so-called angiogenesis [8] and enhances nutrient uptake into cancer cells to satisfy metabolic requirements [9]. In addition, intracellular energetic and biosynthetic pathways are altered to sustain cell proliferation [9]. The Warburg effect is the most well-known example of altered glucose metabolism of cancer cells [10]. Normal cells use glucose to allow for energy production in a form of ATP through the oxidation of its carbon bonds. In contrast, cancer cells use glucose mainly by glycolysis, which produces lactate even in the presence of oxygen, instead of the efficient oxidative phosphorylation [10]. The aerobic glycolysis produces ATP at a fast rate than oxidative phosphorylation does, in spite of low efficiency in ATP yield per molecule of glucose [11]. However, glycolysis in cancer cells is not essential to contribute to ATP production since the average contribution of ATP from glycolysis is only 17% [12]. Currently, it is thought that aerobic glycolysis fulfills needs in cancer cells to support macromolecule biosynthesis, beyond ATP production [13]. High demand of macromolecule biosynthesis in proliferating cancer cells also requires precursors, derived from amino acids such as serine and glycine [14].
SERINE-GLYCINE METABOLISM IN CANCER CELLS
Serine synthesis pathway (SSP)
Cancer cells rapidly consume and utilize serine as an intermediate metabolite [14]. Serine is synthesized through SSP, which begins with the glycolytic intermediate 3-phosphoglycerate (3-PG) (Figure 1). 3-PG is converted to 3-hydroxypyruvate by the action of the enzyme, phosphoglycerate dehydrogenase (PHGDH) and then 3-hydroxypyruvate (3P-pyruvate) takes part in a transamination reaction with glutamate catalyzed by the enzyme, phosphoserine aminotransferase (PSAT1), resulting in production of phosphoserine (3P-serine) and alpha-ketoglutarate. Phosphoserine is dephosphorylated by the action of phosphoserine phosphatase (PSPH) and produces serine [15]. Serine is mutually converted into glycine by serine hydroxymethyltransferases (SHMTs), which have two isoforms; SHMT1 in cytoplasm and SHMT2 in mitochondria [16]. SHMTs bridge between SSP and one-carbon metabolism. One-carbon metabolism involves a complex of metabolic network based on biochemical reactions of folate by the one-carbon groups [171819]. Folate, one of the B vitamins, is reduced by activation of dihydrofolate reductase (DHFR), resulting in the generation of tetrahydrofolate (THF) [1920]. SHMTs, especially SHMT2, transfer a carbon unit from serine to THF to form methylene-THF, initiating the folate cycle. The folate cycle is then coupled to the methionine cycle through methyl-THF (mTHF). Furthermore, the adenylation of methionine produces S-adenosylmethionine (SAM) which functions as a methyl donor for other metabolic pathways that require methyl moieties, including histone, DNA and RNA methylation [2122].
Figure 1
Serine-glycine biosynthesis pathway. Serine is biosynthesized from 3-PG by PHGDH, PSAT1, and PSPH. Biosynthesized serine can be converted into glycine by SHMT1/2. This pathway provides precursors for TCA cycle and antioxidant and purine biosynthesis.
3-PG, 3-phosphoglycerate; PHGDH, phosphoglycerate dehydrogenase; PSAT, phosphoserine aminotransferase; PSPH, phosphoserine phosphatase; SHMT, serine hydroxymethyltransferase; TCA, tricarboxylic acid; THF, tetrahydrofolate; mTHF, methyl-THF.
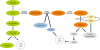
Serine and glycine in cancer cells
Serine and glycine are involved in several biological functions such as glycolysis [23], glutathione (GSH) and nucleotide production [14], and thus are closely associated with proliferation of cancer cell [14]. Serine metabolism and glycolysis are connected by pyruvate kinase M2 isoform (PKM2) [23]. Serine is the allosteric activator of PKM2. PKM2 is inactivated under serine deprivation condition, resulting in an accumulation of upstream glycolytic intermediates. Increased glycolytic intermediates, such as 3-PG, facilitate for metabolic diversion into the SSP [23]. To compensate the low level of glycolysis following PKM2 inhibition, the cells divert a flux of pyruvate into mitochondria to make more energy through tricarboxylic acid (TCA) cycle [23]. Glycine, which is biosynthetically linked with serine, is one of components of GSH, together with glutamate and cysteine. GSH is involved in protection of major cellular components against reactive oxygen species [24] and sensitivity against chemotherapeutic agents such as alkylating agents. Besides, serine and glycine metabolism are involved in nucleotide production, which is highly demanded in cancer cells. Serine is an important precursor of purine and pyrimidine nucleotide biosynthesis in mammalian cells and glycine converted from serine serves as both a carbon and nitrogen source for purine-ring biosynthesis [25].
Serine metabolism is upregulated in many tumors [152627] and PHGDH is the key enzyme of the SSP flux [1528]. The gene encoding PHGDH, located on chromosome 1p12, is mostly amplified in human cancer samples, including breast cancer and melanoma [2930]. Also, colonic tumor tissues have been reported to show significantly increased expression of PHGDH and PSAT, compared to paired normal tissue [31]. In estrogen receptor-negative breast cancer cells, high levels of PHGDH and PSAT were associated with poor prognosis with shorter time to relapse, shorter overall survival time, higher tumor grade, and higher proliferative marker levels [26]. Furthermore, higher expression of PHGDH in glioma and cervical cancer was associated with higher tumor grade [3233].
EPIGENETIC AND SERINE METABOLISM
Epigenetic alteration in cancer
Epigenetics is the study of changes in gene regulation through DNA methylation, histone modification, and non-coding RNA without changes in DNA sequence [3435]. Cytosine in DNA is modified by an addition of a methyl group by DNA methyltransferases (DNMTs) and that produces methylated DNA [36]. DNA methylation is important in cellular differentiation, establishment of genomic imprinting, and regulation of genome stability [37]. Histone modification is another well-known epigenetic mechanism. The N-terminal tails of histones go through various posttranslational covalent modifications such as methylation, acetylation, ubiquitylation, sumoylation, and phosphorylation on specific residues [38]. The modifications activate or repress gene expression by modulating chromatin structure, depending on modified residues and covalently modifying groups.
Aberrant epigenetic changes are considered as a hallmark of cancer [39]. It has emerged that human cancer cells carry epigenetic abnormalities beside genetic alterations and many of the epigenetic alterations play key roles in cancer initiation and progression [404142]. For examples, DNA methyltransferase 3A (DNMT3A), one of major DNMTs, shows somatic mutations in 22% of cases of acute myeloid leukemia (AML) and patients with DNMT3A mutations are associated with shorter overall survival [43]. Similarly, histone methylation has been connected to tumorigenesis. Aberrant patterns of histone methylation by overexpression of histone methyltransferases, EZH2 and EHMT2, are strongly associated with multiple types of cancer [44454647].
Serine metabolism and epigenetic changes
Serine metabolism is linked to one-carbon metabolism, which influences epigenetic patterns through production of SAM. SAM derived from methionine is the major methyl donor in cellular methyl transfer process including DNA/RNA methylation [21]. Although serine is not directly involved in DNA/RNA methylation, serine starvation reduces DNA/RNA methylation levels in cancer cells via lack of regeneration of methionine from homocysteine (Table 1) [48].
Table 1
Summary of the relationships between serine metabolism and epigenetics

| Relationships | Regulator | Results | Ref. |
|---|---|---|---|
| Serine metabolism → epigenetics | Serine | Serine regenerates methionine from homocysteine resulting in contribution to DNA/RNA methylation | [48] |
| LKB1 | LKB1 loss restricts serine metabolism and it causes lower production of SAM leading to lower 5mC level in KRAS mutation cells | [49] | |
| Epigenetics → serine metabolism | EHMT2 | Decrease in SSP enzymes level and serine level in EHMT2 inhibition via decreasing H3K9me1 and increasing H3K9me2 at the promoter of SSP enzymes | [51] |
| KDM4C | Increase in ATF4 mRNA level by lowering H3K9me3 at the promoter of ATF4 leading to increase in SSP enzymes mRNA level | [53] |
The relationship between serine metabolism and DNA methylation has been suggested in the liver kinase B1 (LKB1) deficient model (Table 1) [49]. The cooperation of Kirsten rat sarcoma viral oncogene homolog (KRAS) proto-oncogene (KRAS) activation and LKB1 inactivation results in the synergic oncogenic effect by altering cellular metabolism. In the cells with KRAS activation, LKB1 loss dramatically induces SSP by elevating PSAT1, PSPH, and SHMTs, leading to high SAM production. Furthermore, LKB1 loss increases the expression of methyltransferases, such as DNA methyltransferase 1 (DNMT1) and DNMT3A, resulting in DNA hypermethylation. LKB1 directly regulates 14 kinases related to AMP-activated protein kinase (AMPK), which has a central role in nutrient sensing and reprograming of cell metabolism [50]. LKB1 loss-induced alteration of SSP and DNA methylation takes place in an AMPK-mTOR-dependent manner [49]. The results from the LKB1 model emphasize that serine metabolism-coupled SAM generation directly influences regulation of DNA methylation to support tumorigenesis in cancer cells.
Epigenetic regulation on SSP
There is limited evidence to show that epigenetic modifiers directly regulate SSP. However, a few studies recently suggested several possibilities (Table 1) [5152]. Firstly, EHMT2 regulates SSP [51]. Loss of EHMT2 by activity inhibition or silencing decreases expression of SSP-related genes, PHGDH, PSAT1, PSPH, and SHMT1/2, by reducing mono-methylation and increasing di-methylation at histone H3 lysine 9 (H3K9). In turn, the suppressed SSP reduces the concentration of serine and glycine, leading to cell death. Thus, EHMT2 in cancer provides serine and glycine to support cell proliferation by increasing expression of SSP-related genes. Another epigenetic modifier that regulates SSP is KDM4C. KDM4C is a histone demethylase, targeting histone H3K9. It acts on H3K9 tri-methylation at the promoter of the ATF4 gene and activates the expression [53]. ATF4 is the master regulator of amino acid metabolism including genes encoding SSP enzymes, PHGDH, PSAT1, and PSPH [54]. The results indicate that KDM4C induces SSP by transcriptional upregulation of ATF4 [52].
CONCLUSION
Serine metabolism plays a significant role in proliferating cancer cells by providing precursors for macromolecule biosynthesis. SSP-dependent SAM generation affects epigenetic patterns, in particular changes in methylation on DNA, RNA, and histones, leading to transcriptional changes of oncogenes and tumor suppressor genes. On the other hand, genes encoding SSP enzymes are also directly regulated by epigenetic enzymes, catalyzing acquisition or removal of histone methylation. The evidences suggest that the interactions between SSP and epigenetic regulation contribute to cancer-specific metabolism to sustain rapid proliferation. Further investigations on the regulation of SSP and epigenetic modifiers will shed light upon their potentials as anticancer therapeutic targets.
 XML Download
XML Download