

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Estradiol (17β-estradiol) is a reproductive hormone that is synthesized primarily by the gonads and regulates development and function of the reproductive organs. It also stimulates cell proliferation [22], has anti-inflammatory functions [14], promotes tissue survival [35], and enhances cardiovascular health [7], memory [15], and mood [32]. Estradiol deficiency leads to acne [18], and it is a contributing factor to ovulatory dysfunction [29], menopausal symptoms [31], insulin resistance [17], Alzheimer's disease [27], and bone loss [37]. Estradiol's novel roles in non-reproductive organs have been reported, and efforts have been made to search for the possibilities of estradiol synthesis in extra-gonadal sites. Thus far, bone [34], brain [6], liver [12], adipose tissue [5], skin [3], blood vessels [11], and spleen [33] have been identified as sites of estradiol synthesis; sites in which aromatase (P450arom; CYP19), a key enzyme in estradiol synthesis, is expressed [1].
In the digestive tract, estradiol regulates appetite [21], prevents colon cancer development [836], alleviates syndromes of inflammatory bowel diseases [23], decreases metabolic syndrome in menopausal women [4], and modulates gut microbiome and lymphocyte homeostasis [1019]. Recently, we reported that estradiol is synthesized in murine Peyer's patch (Pp) and mesenteric lymph node (mLN) tissues and has a role in regulating lymphocyte homeostasis in those secondary lymphoid tissues [24]. In this study, to determine whether estrogen synthesis in intestinal lymphoid tissues is conserved in other mammals, we examined the estrogen synthesis capacity of Pp and mLN tissues in pigs.
Materials and Methods
Chemicals and reagents
Minimal essential medium alpha (α-MEM) was purchased from Invitrogen (USA), fetal bovine serum (FBS) from American Type Culture Collection (ATCC; USA), insulin-transferrin-selenium (ITS) from VWR (USA), and androstenedione from Sigma (USA). The sensitive estradiol enzyme-linked immunosorbent assay (ELISA) kit was purchased from DRG International (USA).
Animals
This study was performed in accordance with recommendations in Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. Animal handling was done in accordance with the protocols approved by the University of Illinois at Urbana-Champaign's Institutional Animal Care and Use Committee (protocol No. 16036). Seven naturally farrowed domestic female Yorkshire piglets aged 3 weeks (n = 3) and 12 weeks (n = 4) were obtained from the University of Illinois swine herd and used in this study.
Tissue cultures
To measure estradiol-synthetic capacity, Pp, mLN, and ovary tissues were dissected from individual piglets euthanized by intracardiac administration of sodium pentobarbital (78 mg/kg body weight, Fatal Plus; Vortech Pharmaceuticals, USA), weighed, and cultured overnight in steroid-free media (α-MEM supplemented with 1% ITS [10 ng/mL insulin, 5.5 ng/mL transferrin, 5.5 ng/mL selenium], 5% FBS, and penicillin-streptomycin). Ovary tissue was then washed with fresh media to remove any steroid that might have been released to the culture media. Tissues were further cultured in the absence (vehicle) or presence of androstenedione (200 nM) [24]. Media were collected at 24, 48, and 72 h to measure estradiol content.
Estradiol assays
Estradiol concentrations were measured in blood collected via cardiac puncture, as well as in culture media and tissues (mLNs and gonads). Culture media estradiol content was measured directly, whereas, for blood and tissue samples, total lipids were extracted, following a standard procedure [25], and stored at −80℃ until needed for hormone assay. The concentrations of estradiol in the extracted lipids were measured by using ELISA (EIA-4399; DRG International, Germany) as previously reported [24]. Final estradiol concentrations were expressed by weight per unit volume or by weight of tissue, sera, or culture media (pg/mL or pg/mg). The samples were run in triplicate and had intra- and inter-assay coefficients of variability below 10%. The detection range was 0.1 to 30 pg/mL.
Reverse transcription polymerase chain reaction
For gene expression analysis, mLNs, Pps, and ovaries were collected. Total RNA was extracted, by using TrizolVR solution (Ambion, USA), from porcine tissues (Pps, mLNs, and gonads) and then purified with an RNEasy Kit (Qiagen, USA). Concentration and quality of total RNA were analyzed by using a Nanodrop spectrophotometer (Thermo Scientific, USA) and the samples were then stored at −80℃ until use. Complementary DNA was generated by using M-MLV Reverse Transcriptase (Thermo Scientific) and random primers. The following primers were used for reverse transcription polymerase chain reaction (RT-PCR): 17β-Hsd 5′-AGC CAG AAT ATG TGG CAC CC-3′/5′-CAA CAA GTC CTG ATG GGG CT-3′, Cyp19a1 5′-GGA AAT CCA GAC TGT TGT TG-3′/5′-GCT GGA AGT ACC TGT AAG GA-3′, StAR 5′-GAC TTT GTC GGC TGT-3′/5′-ATC CCT TGA GGT CAA TGC TC-3′, 3β-Hsd 5′-CGT CCT GAC ACA CAA CTC CAA-3′/5′-CCA CGT TGC CGA CGT AGA-3′, CYP17a1 5′-TCC GAG AGG TGC TTC GAT TC-3′/5′-GGC GCT CCT TGA TCT TCA CT-3′, Rpl19 5′-CCT GAA GGT CAA AGG GAA TGT G-3′/5′-GTC TGC CTT CAG CTT GTG GAT-3′. PCR products were run on 2% agarose gels containing ethidium bromide and photographed under UV light. The RT-PCR products were quantified by determining band intensities measured with 1Dscan EX (Scanalytics, USA); Rpl19 was used as the internal control.
Histology and immunocytochemistry
Collected tissues were fixed in 4% paraformaldehyde, embedded in paraffin wax, sectioned at 6 µm thickness, and stained with hematoxylin and eosin. Immunofluorescent labeling of CYP19 and PNAd were performed as previously described [24] by using monoclonal anti-CYP19 (MCA2077S; Bio-Rad Laboratories, USA) or purified rat anti-mouse PNAd carbohydrate epitope (MECA-79; BD Biosciences, USA). After primary antibody incubation overnight, slides were incubated with a secondary biotinylated horse anti-mouse IgG (Vectastain ABC kit; Vector Labs, USA) or rabbit anti-rat IgG (Vectastain ABC kit) and then treated with avidin-biotin complex solution (Vectastain Elite ABC kit; Vector Labs). For color development, 3,′-diaminobenzidine (DAB; Vector Labs) was applied. Slides were then counter-stained with hematoxylin, mounted, and imaged by using an Olympus BX51 microscope (Olympus, Japan).
Statistical analysis
Datasets were first tested for normality and homogeneity of variance and were transformed before statistical analysis. All figures depict non-transformed data. Statistical analyses were performed by applying two-tailed Student's t-test or one-way analysis of variance (ANOVA) followed by Tukey's honest significant difference.
Results
Porcine mesenteric lymph node and Peyer's patch contain and produce estradiol
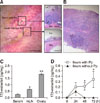
In porcine species, the ileum contains multiple Pps. At three weeks of age, Pps are visibly identifiable by their unique luminal form, which appears as whitish round segments. These segments are follicles that contain densely populated lymphocytes (panel A in Fig. 1; insets). In pigs at this age, mLNs are easily identifiable as they are morphologically distinctive, but the follicles in this organ are less clearly defined than those in Pps (panel B in Fig. 1). The estradiol concentrations of serum, mLN, and ovary were 0.27 pg/mg, 0.66 pg/mg, and 1.84 pg/mg, respectively (panel C in Fig. 1). To see if lymphoid tissues synthesize estradiol, Pps-containing and Pp-free ileal tissue samples were cultured for 72 h, and the estradiol concentrations in the culture media were measured. During the culture period, estradiol concentrations continued to rise in the culture media containing Pps, but not in the media without Pps (panel D in Fig. 1). When the estradiol-synthetic capacity of the Pps was compared with those of mLN and ovary in 12-week-old pigs, the mLN and Pp samples exhibited a robust estradiol-synthetic capacity (Fig. 2). Notably, the estradiol-synthetic capacity of mLN (12.36 pg/mg at 72 h) was comparable to that of ovary (12.45 pg/mg at 72 h). Addition of androstenedione to the tissue cultures increased estradiol production slightly, but not significantly, in all examined tissues.
Porcine mesenteric lymph node and Peyer's patch express mRNAs for steroidogenic enzymes
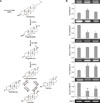
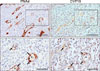
De novo estradiol synthesis in tissue requires the expression of StAR, 3β-Hsd, Cyp17a1, Cyp19a1, and 17β-Hsd genes (panel A in Fig. 3). To determine whether mLNs and Pps synthesize estradiol de novo, mRNA expression levels of steroidogenic genes were examined by performing RT-PCR (panel B in Fig. 3). Both Pps and mLNs expressed mRNAs for all of the steroidogenic genes examined, with minor differences in expression levels. Specifically, 3β-Hsd mRNA expression in both Pps and mLNs were significantly lower than the ovarian expression level (p < 0.01). The expression levels of 17β-Hsd in Pps and mLNs were 2-fold higher than that in ovary (p < 0.01). Pps and mLNs showed lower Cyp19a1 mRNA expression than that in ovary (panel B in Fig. 3). Importantly, immunohistochemical results showed that ileal tissue containing Pps of 12-week-old piglets expresses aromatase protein (CYP19) in endothelial cells of high endothelial venules (HEVs) (Fig. 4).
Discussion
Both mLN and Pp tissues function at the forefront of immune surveillance in the gastrointestinal tract. They recruit, store, and dispatch lymphocytes, which serve as the first respondents to external signals that trigger immune responses [913]. This study aimed to determine whether Pps and mLNs of gilts synthesize estradiol.
If a tissue synthesizes estradiol, the estradiol concentration in that tissue will most likely be higher than the level in circulation. In the three-week-old piglets, the mLN and Pp tissues contained estradiol at higher concentrations than the serum level, indicating an estradiol-synthetic capacity of these two lymphoid tissues. We verified their estradiol-synthetic capacity by culturing them in vitro and measuring estradiol concentration in the tissue culture media. The tissue culture experiment revealed that ileal tissues containing Pps produced estradiol, but the Pp-free ileal tissue did not, indicating that the estradiol synthesis machinery resides within the Pp. This finding is consistent with a previous study showing that Pps, but not Pp-free ileal tissues, synthesize estradiol in mice [24].
Porcine gilts reach puberty as early as 12 weeks of age, at which point the steroidogenic activity of their ovaries begins, thereby producing a large amount of estradiol that triggers a preovulatory luteinizing hormone surge [226]. To compare the estradiol-synthetic capacity of Pps and mLNs with peri-pubertal ovaries, this study used 12-week-old piglets. Surprisingly, our results showed that mLN and ovary tissues had comparable estradiol-synthetic capacity.
Measurement of RNA transcription showed that Pps and mLNs express mRNAs for steroidogenic enzymes related to estradiol precursor production (StAR, 3β-Hsd, and Cyp17a1) and estradiol synthesis (Cyp19a1 and 17β-Hsd). This result indicates that Pps and mLNs may, similar to that in murine counterparts [24], synthesize estradiol de novo, starting from cholesterol (Fig. 3). In support of those results, both the mLN and Pp tissues synthesized estradiol without the addition of androstenedione to the culture media. Of note, 3β-Hsd mRNA expression levels in Pps and mLNs were significantly lower than that in ovarian tissues. Interestingly, 17β-Hsd and Cyp19a1 mRNA expression patterns in ovary were the opposite of the patterns observed in the lymphoid tissues: 17β-Hsd mRNA expression was higher in lymphoid tissues than in ovary, while Cyp19a1 mRNA expression was lower in lymphoid tissues than in ovary. These results suggest that, in Pp and mLN tissues, 17β-HSD may play a significant role in producing estradiol. In the immunohistochemical results, PNAd (a HEV marker) was observed to be localized with CYP19 in the endothelial cells of HEV, suggesting that these cells are the cell types responsible for estradiol synthesis in the examined lymphoid tissues. This result is not surprising because the same cell type was reported to be estrogen-producing in the murine gastrointestinal tract [24].
The roles that estradiol have in the examined lymphoid tissues are yet to be determined. However, estradiol probably has a direct effect on the function and behavior of lymphocytes (cytokine production, differentiation, and proliferation) [16202830]. A homeostatic role of locally synthesized estradiol has been suggested [24], with the implication that dysregulated estradiol synthesis in the gastrointestinal tract may leave the intestine prone to hyperinflammation. Future studies may investigate this possibility and other roles of locally produced estradiol.

 XML Download
XML Download