

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Renal cell carcinoma (RCC) accounts for 3% of all cancers and 90% of all kidney cancers. RCC is also the third most recurrent urologic tumor [12]. At first diagnosis, circa 30% of patients with RCC have metastases. Additionally, 20% of the patients diagnosed with a clinically localized disease will develop metastases, regardless of the use of curative nephrectomy. Therefore, the mean survival of metastatic RCC is only 12 months [345]. Furthermore, limited therapeutic options are currently available. For instance, metastatic RCC is insensitive to chemotherapy, and drugs for targeted therapies and immunotherapies are ineffective in the treatment of this disease [6]. Thus, there is an urgent need to explore novel therapeutic agents against metastatic RCC.
Transient receptor potential melastatin member 7 (TRPM7) is a nonselective cation channel. Furthermore, it is a member of the diverse TRPM subfamily and part of the transient receptor potential (TRP) superfamily [7]. A recent study showed the participation of TRP channels in cell homeostasis, including cell proliferation, differentiation, apoptosis [8]. Specifically, TRPM7 is well known as an essential regulator of Mg2+ homeostasis and a transporter of other cations [910]. Structurally, TRPM7 is comparable to TRPs and possesses six transmembrane domains, which form a hydrophobic central pore or channel. Additionally, TRPM7 transmembrane domains have amino- and carboxyl-terminals (N- and C-terminals) facing the intracellular side of the cell membrane flanking the transmembrane segments [11]. TRPM7 is a bifunctional protein that has the ability to function as an ion channel and a protein kinase, participating in cancer and other diseases. Indeed, some TRP channels have been associated with the growth and development of cancer [12] and recent research has suggested some mechanisms underlying the role of TRPM7 in human cancers. Nonetheless, the functional relationship between TRPM7 and RCC has not been well documented. Moreover, the effect of TRPM7 knockdown on RCC cell migration and invasion and the underlying mechanism have not yet been determined. Hence, we aimed to explore the role of TRPM7 in RCC.
Go to : 
MATERIALS AND METHODS
This study was approved by the Institutional Review Board of Kyungpook National University Chilgok Hospital (approval number: KNUMC 2016-05-021).
1. Cell culture
The ACHN, SN12C, Caki-1, and Caki-2 cell lines were obtained from the Korean Cell Line Bank (Seoul, Korea) and American Type Culture Collection (ATCC, Manassas, VA, USA). All kidney cancer cells were grown in Dulbecco's modified Eagle's medium (DMEM) (Hyclone, Logan, UT, USA) supplemented with penicillin (100 U/mL), streptomycin (100 mg/mL), and 10% fetal bovine serum (FBS) and incubated at 37℃ in a humidified atmosphere containing 5% (v/v) CO2.
2. TRPM7 small interfering RNA (siRNA) and transfection
AccuTarget Predesigned siRNAs specific for human TRPM7 and scramble siRNAs purchased from Bioneer (Daejeon, Korea) were used to knockdown TRPM7 expression. The sequences for human TRPM7 siRNA were Fwd 5′-GUC UUG CCA UGA AAU ACU CUU-3′ and Rev 5′-GAG UAU UUC AUG GCA AGA CUU-3′. For transient transfection, cells were grown to 80% confluence, and Lipofectamine® RNAiMAX (13778-150; Invitrogen, Carlsbad, CA, USA) was used. Knockdown efficiency was determined using Western blot analysis.
3. Reverse transcriptase-polymerase chain reaction (PCR)
Total RNA was isolated using the TRIzol reagent. The Revert Aid First strand cDNA synthesis kit (K1621; Thermo Fisher Scientific, Waltham, MA, USA) was used for reverse transcription. PCR was performed by using a Maxime PCR Premix (25025; iNtRON Biotech, Seongnam, Korea). The cycling conditions were as follows: 95℃ for 2 minutes and 35 cycles at 94℃ for 20 seconds, 56℃ for 40 seconds, and 72℃ for 90 seconds. The primers for TRPM7 were Fwd 5′-TAG CCT TTA GCC ACT GGA C-3′ and Rev 5′-GCA TCT CCT AGA TTT GC-3′. For β-actin, the primers were Fwd 5′-CAT CCT GCG TCT GGA CCT G-3′ and Rev 5′-ATC TCC TTC TGC ATC CTG TC-3′.
4. Western blot analysis
Total cell lysates were prepared in lysis buffer (#9803; Cell Signaling Technology, Beverly, MA, USA) and centrifuged at 14,000 ×g for 10 minutes. Proteins (50 µg) were loaded into a sodium dodecyl sulfate-polyacrylamide gel and transferred onto nitrocellulose membranes for immunoblotting analysis. An anti-β-actin antibody (#4967, rabbit polyclonal, 1:1,000; Cell Signaling Technology) was used as an internal loading control. An anti-TRPM7 antibody (ab85016, mouse monoclonal, 1:1,000) was purchased from Abcam (Cambridge, UK), and the rabbit polyclonal antibodies (1:1,000) against matrix metalloproteinase (MMP)-2 (#4022), MMP-9 (#2270s), Akt (#9272), phospho-Akt (#9271, Ser473), p38 (#9212), phospho-p38 (#9211, Thr180/Tyr182), Src (#2108), phospho-Src (#2105, Tyr527), ERK1/2 (#9102), phospho-ERK1/2 (#9101, Thr202/Tyr204), JNK (#9252), phospho-JNK (#9251, The183/Tyr185) were purchased from Cell Signaling Technology. Immunoreactive protein bands were visualized using a chemiluminescent substrate (Thermo Fisher Scientific).
5. Cell proliferation assay
For the cell viability assay, ACHN and SN12C cells were seeded at 1×105 cells/mL and cultured for 24 hours before transfection with 50 to 100 pmole/µL siRNA for 24 hours. After treatment, 20 µL/well of MTS from a cell proliferation colorimetric assay kit (K300; BioVision, Milpitas, CA, USA) was added, followed by a 2-hour incubation at 37℃ in the dark. Subsequently, the medium was removed, and the formazan precipitate was dissolved in dimethyl sulfoxide (34869; Sigma-Aldrich, St Louis, MO, USA). The absorbance of the formazan product was measured at 490 nm using an enzyme-linked immunosorbent assay (ELISA) reader (BioTek, Winooski, VT, USA).
6. Wound healing assay
For wound healing assay, the surface of cell monolayers in 6-well plates were scratched with a pipette tip. The wounded cells were washed several times with phosphate-buffered saline to eliminate debris. Subsequently, DMEM containing Lipofectamine (25 pmole) and TRPM7 siRNA (50–100 pmole) were added into the scratched wells. The cells were then incubated for 24 hours at 37℃. The initial wound and migration of the cells into the scratched area were photographically monitored and imaged at 0 and 24 hours using an Olympus CKX41 inverted microscope coupled with a digital imaging system.
7. In vitro migration assay
A 24-well Transwell plate system (Costar; Corning Inc., Corning, NY, USA) was used to analyze cell migration. Kidney cancer cells were implanted at a density of 5×104 cells/well onto 8.0 µm Transwell inserts. Inserts were filled with 300 µL of cell suspension, and the lower chamber was filled with 700 µL of DMEM containing 10% FBS. The cells were incubated for 24 hours or 48 hours at 37℃ (5% CO2). Pictures (at 40× magnification) of the membrane were taken in 10 random fields per chamber. After imaging, all Transwell membranes were harvested by incubating the inserts in 100 µL of DMSO for 20 minutes. An ELISA reader (BioTek) was used to detect the absorbance intensity at 595 nm. Each experiment was performed in triplicate.
8. In vitro invasion assay
Invasion assays were performed as previously described. Briefly, 300 µL of cell suspensions (5×104 cells) in DMEM supplemented with 10% FBS were added into Matrigel-coated invasion chambers (8.0-µm, 24-well plates, Costar; Corning Inc.) for 2 hours at 37℃. Photographs were taken, and membranes were harvested by incubating the wells in 100 µL DMSO for 20 minutes. Absorbance was measure at 595 nm on an ELISA reader (BioTek).
9. Inhibitor treatments
Src (ab141987, SKI-606, Bosutinib) and Akt1/2 (#9901, LY294002) inhibitors obtained from Sigma-Aldrich and Abcam, respectively; were used in in vitro migration and invasion assays.
10. Statistical analysis
Data were expressed as mean±standard error. Student's t-test and ANOVA were used to compare groups and determine statistical significance. All statistical analyses were performed using the Statistical Package for the Social Sciences with PASW Statistics ver. 18.0 software (IBM Co., Armonk, NY, USA). A p-value <0.05 was considered statistically significant.
Go to :
RESULTS
1. TRPM7 expression and knockdown
To determine whether TRPM7 is expressed in RCC cell lines, TRPM7 mRNA and protein expression levels were assessed in different RCC cells (Caki-1, Caki-2, SN12C, and ACHN cell lines). As TRPM7 was more highly expressed in ACHN and SN12C cells than in the other cell types, they were selected for knockdown experiments (Fig. 1A). TRPM7 siRNA was used to knockdown its expression, and the efficiency of this knockdown was determined by analysis the protein expression levels of TRPM7 by Western blot. The results show that TRPM7 expression was successfully silenced with TRPM7 siRNA in both ACHN and SN12C RCC cells compared with that in mock and negative control cells (Fig. 1B).
 | Fig. 1(A) Differentially expressed mRNA and protein levels of transient receptor potential melastatin member 7 (TRPM7) in various renal cell carcinoma lines. ACHN and SN12C cells were selected for gene silencing experiments because of the higher expression levels of TRPM7. (B) Small interfering RNA-mediated knockdown of TRPM7 in ACHN and SN12C cell lines analyzed by Western blotting. β-actin was used as an internal loading control. The band is a representative of three independent experiments. M, mock; N, negative control.
|
2. Effect of TRPM7 silencing on RCC cell growth
To assess the effects of TRPM7 gene silencing on RCC cells, we first analyzed cell viability using the MTS assay. After transfection of ACHN and SN12C cells with TRPM7 siRNA at different doses, silencing of TRPM7 had a weak suppressive effect on ACHN cells, but this effect was not significant. Moreover, silencing of TRPM7 had no effect on SN12C cell viability (Fig. 2).
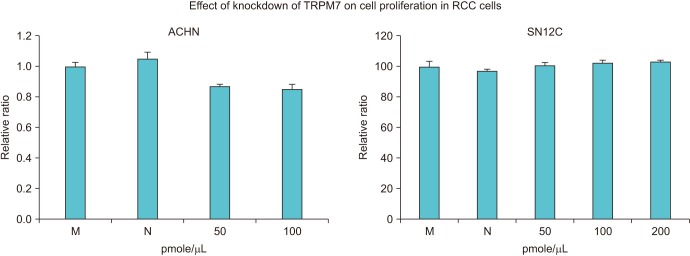 | Fig. 2Effect of the small interfering RNA (siRNA)-mediated knockdown of transient receptor potential melastatin member 7 (TRPM7) on the viability of renal cell carcinoma (RCC) cells. The viability of RCC cells was measured by MTS assay after transfection with TRPM7 siRNA. No significant effect on viability were induced by silencing of TRPM7. M, mock; N, negative control.
|
3. Migration and invasion are suppressed in TRPM7 siRNA-transfected RCC cells
Next, we focused on whether TRPM7 gene silencing influences the migration and invasion ability of RCC cells. Cells that were 90% confluent were scratched with a pipette tip and incubated with either TRPM7 siRNA, mock siRNA, or negative control (transfection reagent alone) for 24 hours. As demonstrated in Fig. 3, a decrease in cell migration to the scratched area was observed following TRPM7 siRNA treatment. Subsequently, the effect of TRPM7 siRNA on the migration of RCC cells was examined by Transwell assays. As shown in Fig. 4A, knockdown of TRPM7 significantly inhibited the migration ability of ACHN and SN12C cells. Furthermore, the effect of TRPM7 siRNA on the metastatic potential of the RCC cells was examined by using an invasion assay. The invasive rates were reduced by TRPM7 siRNA transfection (Fig. 4), suggesting that silencing TRPM7 expression inhibits the metastatic potential of RCC cells.
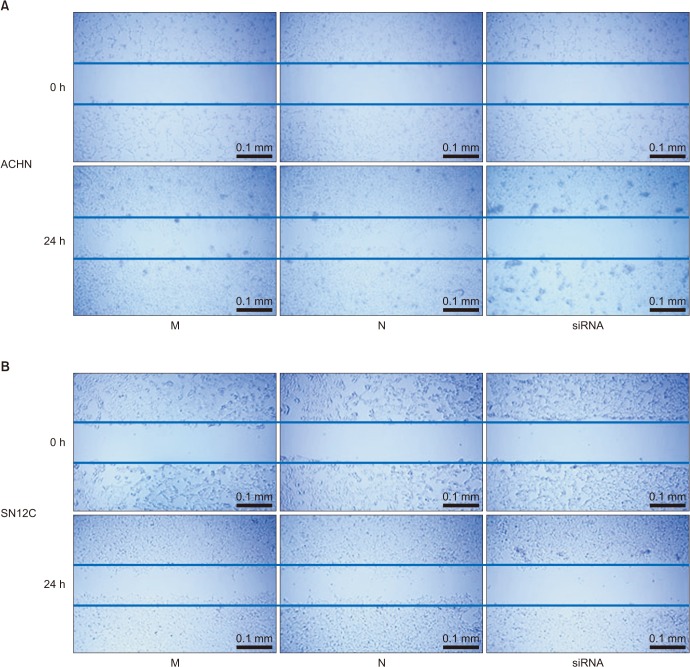 | Fig. 3Effect of transient receptor potential melastatin member 7 (TRPM7) small interfering RNA (siRNA) on cell migration. Wound healing assays reveal a significant decrease in migration of renal cell carcinoma cells after TRPM7 knockdown for 24 hours. (A) ACHN cells. (B) SN12C cells. M, mock; N, negative control.
|
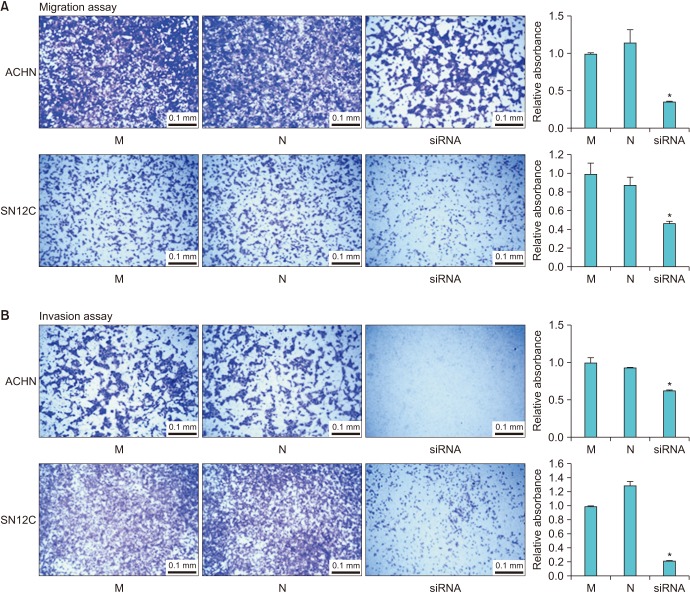 | Fig. 4Effect of small interfering RNA (siRNA)-mediated knockdown of transient receptor potential melastatin member 7 (TRPM7) on the migration and invasion of ACHN and SN12C cells. (A) Effect of TRPM7 siRNA on the migration of renal cell carcinoma (RCC) cells was examined by Transwell assay. The difference in migration was measured as a fold change in absorbance as compared to mock (M) siRNA. The Transwell assay showed that knockdown of TRPM7 significantly inhibited the migration ability of ACHN and SN12C cells. (B) Invasion of RCC cells was investigated by using Matrigel-coated chambers. The number of invading cells was presented as a fold difference relative to M siRNA. The invasion rates were reduced by TRPM7 siRNA transfection, suggesting that silencing of TRPM7 inhibits the metastatic potential of RCC cells. Bar graph shows quantitative analysis of crystal violet extracted from the cells placed under the Transwell. *Significant difference at p<0.05. N, negative control.
|
4. TRPM7 gene silencing leads to a decrease in the phosphorylation Src and Akt
To determine the mechanism of action by which TRPM7 influences the migration and invasion of RCC cells, the phosphorylation of Src, ERK, JNK, p38, and Akt was examined by immunoblotting. After silencing of TRPM7, a considerable decrease in the phosphorylation level of Src was observed compared to that with mock siRNA and negative control (Fig. 5). Similarly, the phosphorylation level of Akt in ACHN RCC cells was reduced considerably after silencing of TRPM7 compared to that with mock siRNA (Fig. 5). To confirm that TRPM7 knockdown-mediated inhibition of migration and invasion is indeed linked to Src and Akt phosphorylation, the cells were treated with the Src inhibitor SKI-606 and the Akt inhibitor LY294002. Fig. 6A shows that treatment with Akt or Src inhibitor reduced the migration ability of ACHN cells in wound healing assays. Moreover, the Src inhibitor decreased the migration of SN12C cells to the scratched area (Fig. 6B). In Fig. 7A, the Transwell assay showed that the Src inhibitor significantly inhibited the migration ability of ACHN and SN12C cells. In ACHN cells, the Akt inhibitor considerably suppressed cell migration as indicated by the Transwell assay (Fig. 7A). The effect of Src and Akt inhibition on RCC invasion was also examined using an invasion assay. Invasion rates were reduced by inhibition of both Src and Akt, suggesting that TRPM7 silencing inhibits the metastatic potential of RCC cells via inactivation of the Src and Akt signaling pathways (Fig. 7B).
 | Fig. 5Effect of transient receptor potential melastatin member 7 (TRPM7) down-regulation on Akt and Src phosphorylation in renal cell carcinoma cells. (A) In ACHN cells, the phosphorylation levels of Akt and Src were significantly reduced by the small interfering RNA (siRNA)-mediated knockdown of TRPM7. (B) Western blot images show decreased phosphorylation of Src in SN12C cells following transfection with TRPM7 siRNA. M, mock; N, negative control.
|
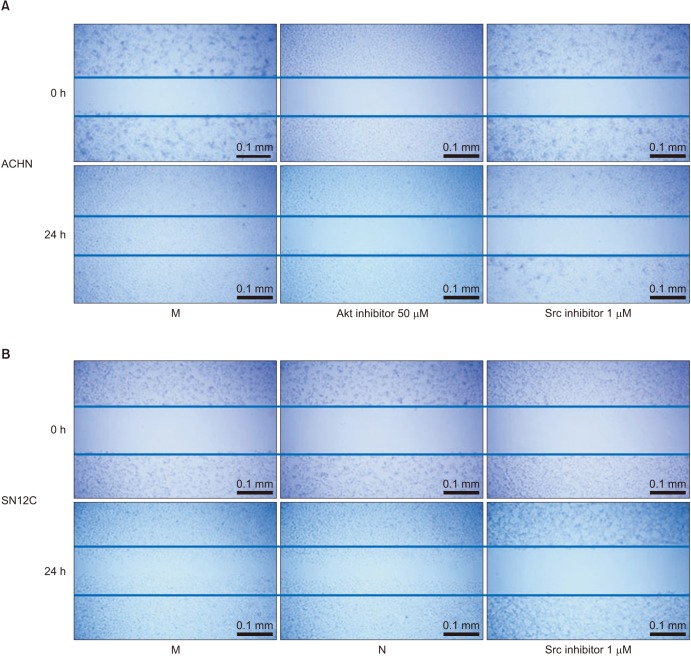 | Fig. 6Effect of Akt and Src inhibition on cell migration. (A) Treatment with Akt or Src inhibitor of ACHN cells reduced cell migration in wound healing assays. (B) In SN12C cells, the Src inhibitor decreased the cell migration to the scratched area. M, mock; N, negative control.
|
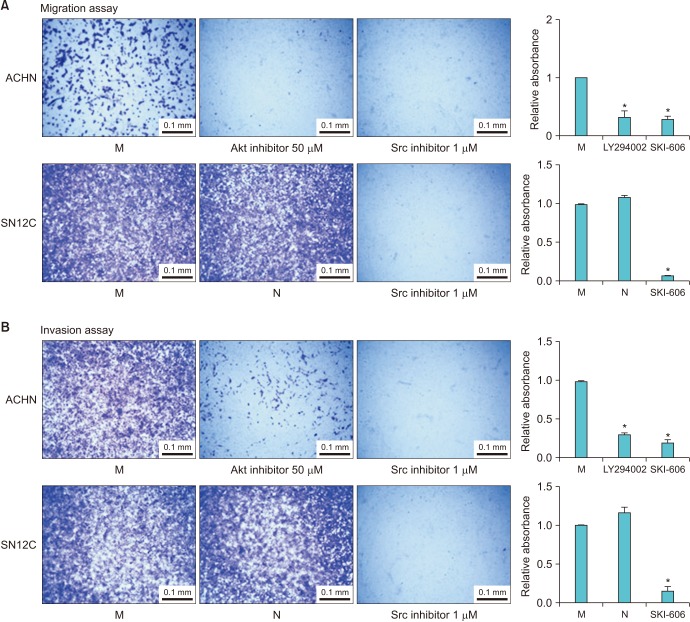 | Fig. 7Effect of Akt and Src inhibition on cell migration and invasion. (A) Transwell assays show that Src inhibition significantly inhibited the migration ability of ACHN and SN12C cells. In ACHN cells, the Akt inhibitor was effective in inhibiting cell migration. (B) The effect of Src and Akt inhibition on the invasion capabilities of renal cell carcinoma (RCC) cells was also examined. Src and Akt inhibitors reduced RCC invasion rates, suggesting that transient receptor potential melastatin member 7 silencing inhibits the metastatic potential of RCC cells via inactivation of the Src and Akt signaling pathways. Bar graph shows quantitative analysis of crystal violet extracted from the cells placed under the Transwell. *Significant difference at p<0.05. M, mock; N, negative control.
|
5. TRPM7 knockdown in RCC cells is not linked to MMP-2 and MMP-9 expression
Recently, a study has reported that MMP-9 expression is associated with migration, invasion, and aggressiveness of tumor cells [13]. Thus, we examined whether TRPM7 knockdown inhibits the expression of MMP-2 and MMP-9 in RCC cells. MMP-2 and MMP-9 were not reduced by TRPM7 siRNA transfection of RCC cells (Fig. 8).
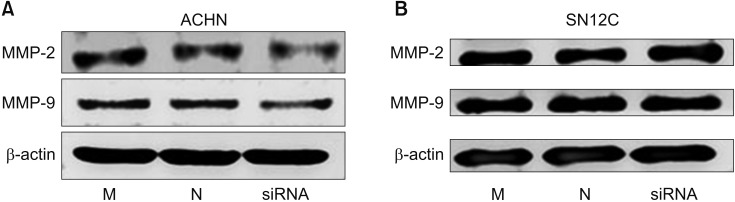 | Fig. 8Effect of transient receptor potential melastatin member 7 (TRPM7) down-regulation on matrix metalloproteinase (MMP)-2 and MMP-9 phosphorylation in ACHN (A) and SN12C renal cell carcinoma (RCC) cells (B). MMP-2 and MMP-9 were not reduced by TRPM7 small interfering RNA (siRNA) transfection of RCC cells. M, mock; N, negative control.
|
Go to :
DISCUSSION
To date, kidney cancer remains the ninth most prevalent malignancy among men, with an estimated 1,131 deaths in Korea in 2017 [14]. Kidney cancer progression varies from slow to highly aggressive. When kidney cancer metastasizes, the mean survival of patients is limited. Given the heterogeneity of RCC, the vigorous study of the underlying molecular mechanisms is a sensible approach [15]. Moreover, metastasis of RCC is a significant obstacle for the systemic treatment of this disease [1617]. Metastasis, the migration of tumor cells from their original site to distant organs, is an intricate multistep mechanism, which involves complex cell adhesion, invasion, and migration [18]. Despite the significance of metastasis to RCC survival and morbidity, little is known on the cellular and molecular mechanisms mediating RCC metastasis.
TRPM7, a well-known member of the TRP protein family, is an ion channel and protein kinase that is ubiquitously expressed in several normal tissues [9]. Recently, TRPM7 has been implicated in cancer metastasis and carcinogenesis [19]. In this study, the evidence collected from wound healing and Transwell assays distinctly shows reduced migration and invasion in ACHN and SN12C RCC cells after the silencing of TRPM7 (Figs. 3, 4). However, cell proliferation remained unaffected (Fig. 2). These results suggest that TRPM7 plays an important role in the invasion phenotype of ACHN and SN12C RCC cells. A recent study showed that depressing the function of TRPM7 is known to inhibit the migration and invasion of MDA-MB-435 breast cancer cells, which is consistent with our data [19]. Besides, up-regulation of TRPM7 augments the migration of A549 lung cancer cells and vascular smooth muscle cells [2021].
Although there is a substantial amount of evidence that indicates that TRPM7 is required for controlling cell migration and invasion, the signaling pathways that mediate its action remain largely undiscovered. In the current study, we investigated the expression and activation of various signaling molecules to examine the pathways involved in RCC migration and invasion that are dependent on TRPM7. Akt, the main signal transducer of the phosphoinositide 3-kinase (PI3K) pathway, promotes the progression of tumor cells by inhibiting apoptosis, promoting cell proliferation, and regulating cell migration and invasion [22]. In RCC, the PI3K/Akt pathway is moderately mutated but highly activated. A large number of patients with RCC were recently involved in an extensive integrated analysis of the PI3K/Akt pathway. The results reiterated the crucial role of the PI3K/Akt pathway in this cancer [23]. Indeed, PI3K pathway inhibitors of the rapalog family have been approved for use in RCC therapy [24]. Our data show that activity of Akt was altered, when the migration and invasion capacity of ACHN cells was decreased by the silencing of TRPM7 (Fig. 5). Moreover, the Akt1/2 inhibitor LY294002 further reinforced the role of Akt in cell migration and invasion of RCC (Figs. 6, 7). Therefore, the conjecture that the TRPM7-mediated migration and invasion of ACHN RCC cells was associated with the Akt signaling pathway is justified. Nevertheless, these results were not apparent in SN12C cells.
Cancer metastasis generally involves the complex coordination of signal transduction pathways. As these signaling pathways control cancer cell migration, invasion, and the reestablishment of tumors, the potential role of Src and mitogen-activated protein kinase (MAPK) in RCC metastasis was considered and studied. These two signaling pathways are known to regulate crucial occurrences in metastasis, such as cancer cell invasion and migration [2526]. Indeed, a previous study indicated that a member of the Src kinase family is associated with a poor prognosis in patients with RCC and could be a potential therapeutic target for RCC [27]. Moreover, a recent study identified that the activation of p38-MAPK signaling was associated with RCC metastasis [28]. Our data demonstrated that the siRNA-mediated silencing of TRPM7 led to a considerable reduction in the migration and invasion capacity of ACHN and SN12C cells and concomitant inactivation of Src (Fig. 5). This suggests that TRPM7 alteration of the migration and invasion of RCC cells involves the Src signaling pathway. To support these findings, ACHN and SN12C cells were treated with the Src inhibitor SKI-606 (Figs. 6, 7). The results indicated that the inhibition of Src reduced the migration and invasion abilities of these cells. Similar results were seen in a previous report in which Src inhibition promoted tumor cell inhibition in RCC [29]. Epithelial-mesenchymal transition in RCC cells is known to involve both the Src and Akt signaling pathways [30]. Notwithstanding, our data indicated that when the migration and invasion capability of RCC cells was reduced by silencing of TRPM7, the activity of MAPKs, including p38, ERK1/2, and JNK1/2, was not affected.
The current study has some limitations. Overexpression experiments should be performed to identify the exact role of TRPM7 in RCC cells. Moreover, TRPM7 expression in human RCC tissues should be measured. To elucidate the precise effects of TRPM7 in RCC, further studies such as expression studies in human tissues are needed.
Go to :
CONCLUSIONS
To the best of our knowledge, this is the first report to address TRPM7 function and mechanism of action in RCC. We provide compelling evidence that TRPM7 channels play an important role in the migration and invasion of cancer cells in human RCC. There is also significant evidence that the signaling route is through the Akt and Src pathways. In conclusion, TRPM7 down-regulation reduced the migration and invasion activity of RCC cells via inactivation of Src and Akt signaling pathways.
Go to :
 XML Download
XML Download