

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Atopic dermatitis (AD) is a chronic inflammatory skin condition with a drastic impact on pediatric health. The multifactorial pathogenesis of AD is still somewhat of an enigma. Its development involves multiple potential genes, an altered innate and adaptive immune response, and epidermal epithelial dysfunction and is influenced by several environmental risk factors.12 The increased prevalence of AD, particularly in industrialized regions, has been hypothesized to be due to the excessive hygiene accompanying the Western lifestyle reduces exposure of the host's immune system to the education provided by beneficial microbes.34 In particular, infancy and early childhood have been identified as important and vulnerable periods in the development of the gut microbiome, which shapes an individual's disposition to AD.5 This is exemplified by germ-free mice, which lack an appropriately developed immune system and show mucosal alterations, both of which can be restored through colonization with the gut microbiome.6 In this ever-expanding field, researchers are now investigating how the local microbiome influences immunity at distal sites, particularly how the gut microbiome influences other organs, such as the lung, brain, and skin. This has led to the coining of terms, such as the “gut-brain axis” and “gut-lung axis.”
Research into the effects of probiotics has revealed how gut microbiome can affect other organs. Several studies have demonstrated that probiotics can influence the composition of the gut microbiome and may even exert immunomodulatory effects.78 In response to these promising results, the possible application of probiotics to the therapeutic management of allergic diseases has been investigated in many studies.910 In particular, a considerable body of literature has been published on the effects of probiotics in patients with AD.11 More recently, interest in dissecting the gut-skin axis has been revived by the finding that the consumption of certain lactobacilli by mice can markedly change their overall skin phenotypes.12 The mechanisms by which the gut microbiome affects the immune responses in the skin, and vice versa, are being uncovered, but many questions remain.
In this review article, we summarize the recent findings on the gut and skin microbiome, highlighting the roles of major commensals in modulating skin and systemic immunity in AD.
Go to : 
DOES THE GUT MICROBIOME HAVE AN IMPACT ON AD?
Gut microbiome and AD
The gut microbiome might play a crucial role in the development of AD by regulating immune system maturation through cross-talk between the microbiome and the host, especially in early life.1314 Alterations in the gut microbiome affect the immune system balance via the production of metabolites, which can cause an inflamed microenvironment in the presence of specific microbiome in the gut.15 The established dysbiosis of the gut microbiome combined with the immune system imbalance persists into adulthood and thereby contributes to the natural courses of diseases, such as AD.16
Various environmental factors, such as stress, diet, and pollutants, affect microbial composition and profiling, particularly in early life.16 In combination with these diverse factors, the gut microbiome contributes to the development and natural course of AD. Previous studies have found that the gut microbiome in early life is associated with age of onset, severity, remission, flares, and even phenotype of AD (e.g., atopic vs non-atopic AD) (Table 1). Although some studies suggested that gut microbiome diversity is inversely related to the development of AD,17 the association between gut microbiome diversity and AD development remains contradictory.171819 The development of AD might be promoted not only by gut microbiome diversity in itself, but also by interactions between specific gut microbiome, established immune systems, and harmonization of the gut microbiome and the host.
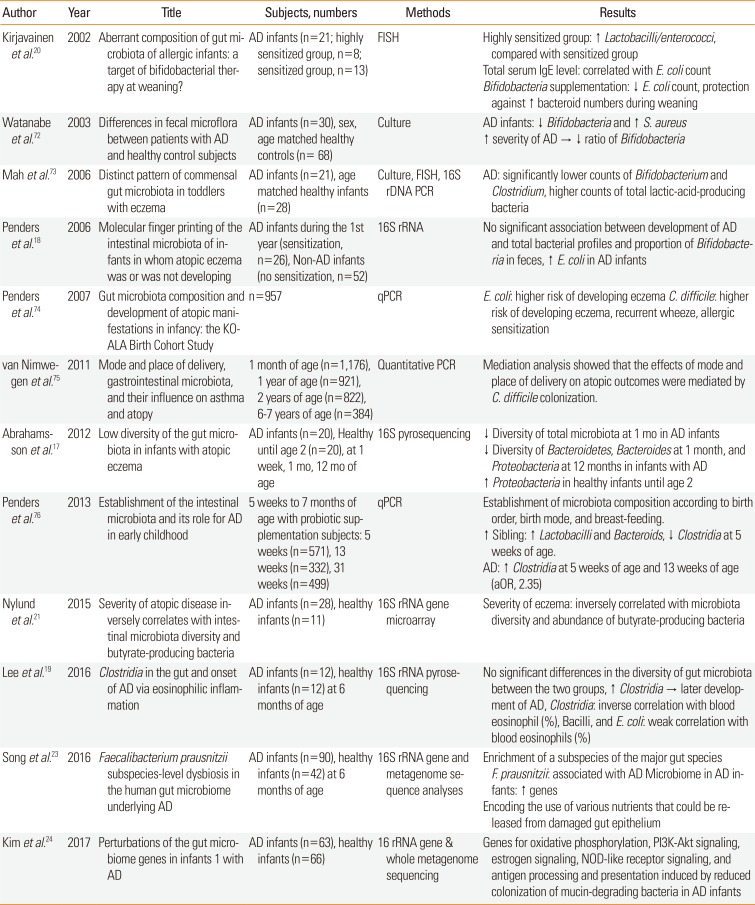
Table 1
Summary of the previous studies of the gut microbiome in AD
| Author | Year | Title | Subjects, numbers | Methods | Results |
|---|---|---|---|---|---|
| Kirjavainen et al.20 | 2002 | Aberrant composition of gut microbiota of allergic infants: a target of bifidobacterial therapy at weaning? | AD infants (n=21; highly sensitized group, n=8; sensitized group, n=13) | FISH | Highly sensitized group: ↑ Lactobacilli/enterococci, compared with sensitized group |
| Total serum IgE level: correlated with E. coli count | |||||
| Bifidobacteria supplementation: ↓ E. coli count, protection against ↑ bacteroid numbers during weaning | |||||
| Watanabe et al.72 | 2003 | Differences in fecal microflora between patients with AD and healthy control subjects | AD infants (n=30), sex, age matched healthy controls (n= 68) | Culture | AD infants: ↓ Bifidobacteria and ↑ S. aureus ↑ severity of AD → ↓ ratio of Bifidobacteria |
| Mah et al.73 | 2006 | Distinct pattern of commensal gut microbiota in toddlers with eczema | AD infants (n=21), age matched healthy infants (n=28) | Culture, FISH, 16S rDNA PCR | AD: significantly lower counts of Bifidobacterium and Clostridium, higher counts of total lactic-acid-producing bacteria |
| Penders et al.18 | 2006 | Molecular finger printing of the intestinal microbiota of infants in whom atopic eczema was or was not developing | AD infants during the 1st year (sensitization, n=26), Non-AD infants (no sensitization, n=52) | 16S rRNA | No significant association between development of AD and total bacterial profiles and proportion of Bifidobacteria in feces, ↑ E. coli in AD infants |
| Penders et al.74 | 2007 | Gut microbiota composition and development of atopic manifestations in infancy: the KOALA Birth Cohort Study | n=957 | qPCR | E. coli: higher risk of developing eczema C. difficile: higher risk of developing eczema, recurrent wheeze, allergic sensitization |
| van Nimwe-gen et al.75 | 2011 | Mode and place of delivery, gastrointestinal microbiota, and their influence on asthma and atopy | 1 month of age (n=1,176), 1 year of age (n=921), 2 years of age (n=822), 6-7 years of age (n=384) | Quantitative PCR | Mediation analysis showed that the effects of mode and place of delivery on atopic outcomes were mediated by C. difficile colonization. |
| Abrahamsson et al.17 | 2012 | Low diversity of the gut microbiota in infants with atopic eczema | AD infants (n=20), Healthy until age 2 (n=20), at 1 week, 1 mo, 12 mo of age | 16S pyrosequencing | ↓ Diversity of total microbiota at 1 mo in AD infants |
| ↓ Diversity of Bacteroidetes, Bacteroides at 1 month, and Proteobacteria at 12 months in infants with AD | |||||
| ↑ Proteobacteria in healthy infants until age 2 | |||||
| Penders et al.76 | 2013 | Establishment of the intestinal microbiota and its role for AD in early childhood | 5 weeks to 7 months of age with probiotic supplementation subjects: 5 weeks (n=571), 13 weeks (n=332), 31 weeks (n=499) | qPCR | Establishment of microbiota composition according to birth order, birth mode, and breast-feeding. |
| ↑ Sibling: ↑ Lactobacilli and Bacteroids, ↓ Clostridia at 5 weeks of age. | |||||
| AD: ↑ Clostridia at 5 weeks of age and 13 weeks of age (aOR, 2.35) | |||||
| Nylund et al.21 | 2015 | Severity of atopic disease inversely correlates with intestinal microbiota diversity and butyrate-producing bacteria | AD infants (n=28), healthy infants (n=11) | 16S rRNA gene microarray | Severity of eczema: inversely correlated with microbiota diversity and abundance of butyrate-producing bacteria |
| Lee et al.19 | 2016 | Clostridia in the gut and onset of AD via eosinophilic inflammation | AD infants (n=12), healthy infants (n=12) at 6 months of age | 16S rRNA pyrosequencing | No significant differences in the diversity of gut microbiota between the two groups, ↑ Clostridia → later development of AD, Clostridia: inverse correlation with blood eosinophil (%), Bacilli, and E. coli: weak correlation with blood eosinophils (%) |
| Song et al.23 | 2016 | Faecalibacterium prausnitzii subspecies-level dysbiosis in the human gut microbiome underlying AD | AD infants (n=90), healthy infants (n=42) at 6 months of age | 16S rRNA gene and metagenome sequence analyses | Enrichment of a subspecies of the major gut species F. prausnitzii: associated with AD Microbiome in AD infants: ↑ genes |
| Encoding the use of various nutrients that could be released from damaged gut epithelium | |||||
| Kim et al.24 | 2017 | Perturbations of the gut microbiome genes in infants 1 with AD | AD infants (n=63), healthy infants (n=66) | 16 rRNA gene & whole metagenome sequencing | Genes for oxidative phosphorylation, PI3K-Akt signaling, estrogen signaling, NOD-like receptor signaling, and antigen processing and presentation induced by reduced colonization of mucin-degrading bacteria in AD infants |
AD, atopic dermatitis; FISH, fluorescence in situ hybridization; E. coli, Escherichia coli; IgE, immunoglobulin E; S. aureus, Staphylococcus aureus; rDNA, recombinant DNA; rRNA, ribosomal ribonucleic acid; qPCR, quantitative polymerase chain reaction; aOR, adjusted odds ratio; NOD, nucleotide-binding oligomerization domain.
![]()
In patients with AD, the proportion of Clostridia, Clostridium difficile, Escherichia coli, and Staphylococcus aureus (S. aureus) in the gut microbiome is higher than in healthy controls, whereas that of Bifidobacteria, Bacteroidetes, and Bacteroides is decreased.1718192021 Butyrate-producing bacteria, such as Coprococcus eutactus, are increased in infants with milder AD or healthy infants than in those with severe AD.21 Short-chain fatty acids (SCFAs), including butyrate, propionate, and acetate, have antiinflammatory and immunomodulatory effects, regardless of their interaction with the gut epithelium barrier.22 In addition, the gut microbiome involved in the SCFA pathway was identified to be present at a higher proportion in patients with AD.23 In association with the mechanisms underlying the development of AD, Clostridia and Escherichia coli in the intestine might be associated with AD via eosinophilic inflammation.19
In another study, the presence of specific gut microbiome, such as Akkermansia muciniphila and Ruminococcus gnavus, was associated with alterations in functional genes, which affected the development of the host immune system.23 It is still unclear whether compositional changes in the gut microbiome precede the development of AD, and thereby shift the immune system and disrupt the gut epithelial barrier, making it amenable to the development of AD, or how the pathogenic role of the established gut microbiome contributes to the development of AD. Gut microbiome and its metabolites might manipulate the local immune responses as well as those of the systemic immune system.1923
One of the effective therapeutic options for AD involves probiotics, although previous studies on the effects of probiotics in the treatment and prevention of the development of AD failed to draw any firm conclusions.25 However, the restoration of gut microbiome dysbiosis can be considered a therapeutic target for AD from the aspect of the harmonization of the gut microbiome with the subsequent immune responses. Both the independent effect of each environmental factor and the combination of the diverse factors involved might shape the immune system, so that it is liable to develop AD. Therefore, studies of the combined effects of the diverse environmental factors and dynamics of the gut microbiome might be more useful to evaluate the impact of gut microbiome in the development of AD.
Go to :
HOW DO GUT MICROBIOME IMPACT AD?
Immunologic pathway
AD is a common clinical manifestation that involves 2 major biological pathways: barrier dysfunction and immune response. AD is associated with a T helper (Th) 1/Th2 imbalance, which induces secretion of Th2 cytokines, such as interleukin (IL)-4, IL-5, and IL-13, and leads to higher production of immunoglobulin E (IgE) and increased binding of S. aureus to AD skin.26
The gut microbiome has been studied as an important contributing factor to the immunologic pathway of AD via probiotics. Orally administered probiotics could interact with gastrointestinal mucosa and gut-associated lymphoid tissue (GALT), where more than 70% of immune cells are located.7 Probiotics interact with epithelial cells, mucosal dendritic cells (DCs), and macrophages through diverse ways. Depending on the probiotic strain, they can either induce immune activation signaling by producing IL-12, IL-18, and tumor necrosis factor (TNF)-α or trigger tolerance signaling by stimulating anti-inflammatory cytokines, such as IL-10 and TGF-β.827 In the IL-10/TGF-β-enriched cytokine milieu, DCs and macrophages can enhance the generation of the induced regulatory T (Treg) cells that play key roles in maintaining peripheral immune tolerance by balancing the ratio of effector and Treg cells. Apart from probiotics, alterations in the gut microbiome might affect the development of host immune cell function through differences in gut microbiome genes, particularly in infants with AD.24
S. aureus is the most common pathogen grown from AD skin, and AD flares are associated with S. aureus superantigens. However, a recent birth cohort study showed that gut colonization by S. aureus strains carrying a certain combination of superantigen and adhesion genes was negatively associated with the subsequent development of AD in infancy.28 Such strains may stimulate and promote the maturation of the infant immune system. Even though skin S. aureus may aggravate already established AD, this does not preclude the possibility that commensal mucosal colonization by gut S. aureus prior to the “atopic march” could have a protective effect through the broad immune stimulation afforded by this bacterium.
Studies in humans also indicate the ability of the gut microbiome to enhance skin health outside the context of AD. In a human study, Lactobacillus paracasei supplementation reduced skin sensitivity and transepidermal water loss in healthy adults.29 The authors attributed these effects to an increase in circulating TGF-β because this cytokine has been shown to affect barrier integrity.30 Thus, all of these studies support the concept that the skin and the gut are linked via modulation of the immune environment through the microbiome.
Go to :
Metabolite pathway
SCFAs produced by the gut microbiome, such as Akkermansia muciniphila,31 play important roles in inflammatory diseases, such as AD, which might explain the association between dietary feeding, microbiome, and the skin immune system.2332 Oral administration of metabolites can affect skin disease by influencing their anti-allergic and anti-inflammatory effects.33 In a mouse study, linoleic acid and 10-hydroxy-cis-12-octadecenoic acid alleviated AD and modulated the gut microbiome. In another study, administration of probiotic Bifidobacterium animalis subsp. lactis (LKM512) increased the levels of the metabolite kynurenic acid, which reduced scratching behavior in AD mice.34
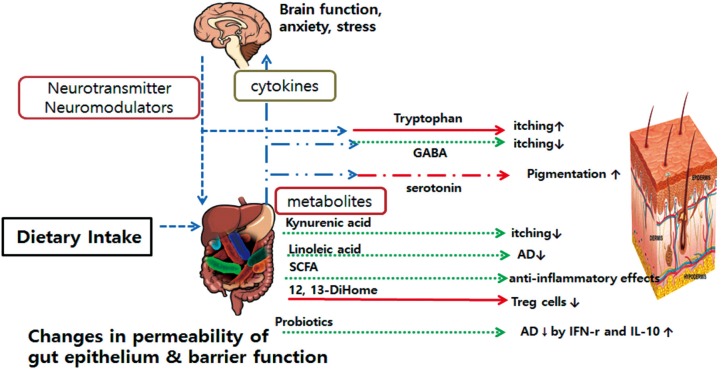
A recently published report showed 3 different states of neonatal gut microbiome (NGM) and metabolite function in early allergic sensitization.35 The so-called NGM3 state in that study was related to multiple allergic sensitizations in asthma and lower relative abundance of Bifidobacterium, Akkermansia, and Facalibacterium. In addition, the metabolite 12,13-dihydroxy-9Z-octadecenoic acid (12,13-DiHome), which has proinflammatory immunomodulatory effects in vitro, was enriched in NGM3. Moreover, 12, 13-DiHome was increased in vernix caseosa as a protective layer in human skin.36 The diverse evidence may support the existence of metabolite-mediated gut-skin axis communications (Figure).
Go to :
Neuroendocrine pathway
The emerging data support the presence of a gut-skin axis that is mediated by neuroendocrine molecules produced by the gut microbiome.3738 The data suggest that compositional and proportional differences in the gut microbiome are linked to the generation of diverse favorable neurotransmitters and neuromodulators, which are associated with the degree of AD symptoms. They can also affect skin barrier dysfunction and immune system dysregulation, which are the key pathophysiologies in the development of AD.
The gut microbiome can modulate the gut-skin axis through direct and indirect pathways.39 Tryptophan produced by the gut microbiome causes an itching sensation in the skin,37 whereas Lactobacillus species and Bifidobacterium species can produce γ-aminobutyric acid (GABA), which inhibits skin itch.3740
Escherichia species and Enterococcus species can produce serotonin, which is involved in skin pigmentation.3841
Indirectly, the gut microbiome can modulate cytokine levels in the bloodstream and thereby affect brain function, anxiety, and stress.39 Cortisol, usually released under stress conditions, can change gut epithelium permeability and barrier function by altering the composition of the gut microbiome.38 This also alters the levels of circulating neuroendocrine molecules, such as tryptamine, trimethylamine, and serotonin, and thereby modifies the skin barrier and skin inflammation.3742 These neuroendocrine molecules might be considered future therapeutics for AD (Figure).
Go to :
HOW DOES SKIN IMMUNITY IMPACT ON SYSTEMIC IMMUNITY AND AD?
Skin microbiome and AD
Chronic inflammation in the skin, such as in AD, exacerbates skin barrier impairment and is associated with abnormalities in the functions of skin barrier-associated genes.434445 Skin microbiomes can influence the skin barrier by controlling ecological factors, such as humidity, temperature, pH, and lipid content.46 Previous research has shown that a compositional change in the skin microbiome influences the development and degree of symptoms of AD via allergic inflammation.47 Dysbiosis of the skin commensal microbiome may contribute to the disruption of immune homeostasis in the skin and promote the development of skin diseases, such as AD.48 Previous studies have found that the skin microbiome is associated with age, lesion of AD, and bacterial composition rate of AD lesion (Table 2). Therefore, determination of whether the skin microbiome plays a role in the initiation of AD could help reduce its development.
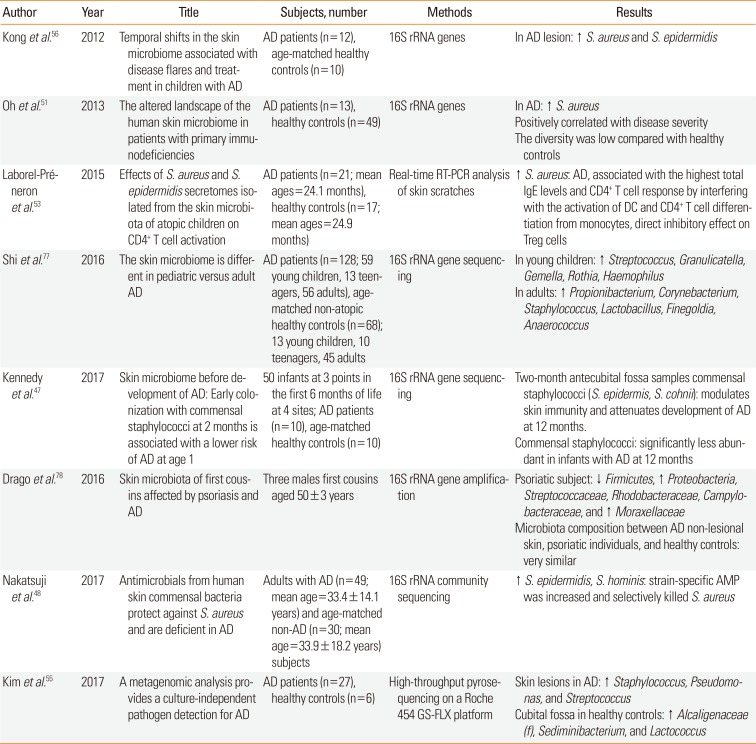
Table 2
Summary of the previous studies on the skin microbiome in AD
| Author | Year | Title | Subjects, number | Methods | Results |
|---|---|---|---|---|---|
| Kong et al.56 | 2012 | Temporal shifts in the skin microbiome associated with disease flares and treatment in children with AD | AD patients (n=12), age-matched healthy controls (n=10) | 16S rRNA genes | In AD lesion: ↑ S. aureus and S. epidermidis |
| Oh et al.51 | 2013 | The altered landscape of the human skin microbiome in patients with primary immunodeficiencies | AD patients (n=13), healthy controls (n=49) | 16S rRNA genes | In AD: ↑ S. aureus |
| Positively correlated with disease severity | |||||
| The diversity was low compared with healthy controls | |||||
| Laborel-Préneron et al.53 | 2015 | Effects of S. aureus and S. epidermidis secretomes isolated from the skin microbiota of atopic children on CD4+ T cell activation | AD patients (n=21; mean ages=24.1 months), healthy controls (n=17; mean ages=24.9 months) | Real-time RT-PCR analysis of skin scratches | ↑ S. aureus: AD, associated with the highest total IgE levels and CD4+ T cell response by interfering with the activation of DC and CD4+ T cell differentiation from monocytes, direct inhibitory effect on Treg cells |
| Shi et al.77 | 2016 | The skin microbiome is different in pediatric versus adult AD | AD patients (n=128; 59 young children, 13 teenagers, 56 adults), age-matched non-atopic healthy controls (n=68); 13 young children, 10 teenagers, 45 adults | 16S rRNA gene sequencing | In young children: ↑ Streptococcus, Granulicatella, Gemella, Rothia, Haemophilus |
| In adults: ↑ Propionibacterium, Corynebacterium, Staphylococcus, Lactobacillus, Finegoldia, Anaerococcus | |||||
| Kennedy et al.47 | 2017 | Skin microbiome before development of AD: Early colonization with commensal staphylococci at 2 months is associated with a lower risk of AD at age 1 | 50 infants at 3 points in the first 6 months of life at 4 sites; AD patients (n=10), age-matched healthy controls (n=10) | 16S rRNA gene sequencing | Two-month antecubital fossa samples commensal staphylococci (S. epidermis, S. cohnii): modulates skin immunity and attenuates development of AD at 12 months. |
| Commensal staphylococci: significantly less abundant in infants with AD at 12 months | |||||
| Drago et al.78 | 2016 | Skin microbiota of first cousins affected by psoriasis and AD | Three males first cousins aged 50±3 years | 16S rRNA gene amplification | Psoriatic subject: ↓ Firmicutes, ↑ Proteobacteria, Streptococcaceae, Rhodobacteraceae, Campylobacteraceae, and ↑ Moraxellaceae |
| Microbiota composition between AD non-lesional skin, psoriatic individuals, and healthy controls: very similar | |||||
| Nakatsuji et al.48 | 2017 | Antimicrobials from human skin commensal bacteria protect against S. aureus and are deficient in AD | Adults with AD (n=49; mean age=33.4±14.1 years) and age-matched non-AD (n=30; mean age=33.9±18.2 years) subjects | 16S rRNA community sequencing | ↑ S. epidermidis, S. hominis: strain-specific AMP was increased and selectively killed S. aureus |
| Kim et al.55 | 2017 | A metagenomic analysis provides a culture-independent pathogen detection for AD | AD patients (n=27), healthy controls (n=6) | High-throughput pyrosequencing on a Roche 454 GS-FLX platform | Skin lesions in AD: ↑ Staphylococcus, Pseudomonas, and Streptococcus |
| Cubital fossa in healthy controls: ↑ Alcaligenaceae (f), Sediminibacterium, and Lactococcus |
AD, atopic dermatitis; rRNA, ribosomal ribonucleic acid; S. aureus, Staphylococcus aureus; S. epidermidis, Staphylococcus epidermidis; RT-PCR, reverse transcription polymerase chain reaction; IgE, immunoglobulin E; DC, dendritic cell; Treg, regulatory T; S. cohnii, Staphylococcus cohnii; S. hominis, Staphylococcus hominis.
![]()
In general, the skin microbiome comprises up to 107 microorganisms per cm2, primarily Propionibacteria, Streptococcus, Staphylococcus, and Corynebacterium.49 Above all, the results of many studies have shown that AD patients have a higher proportion of S. aureus in the skin microbiome compared with healthy controls.485051 In AD patients frequently inflamed with S. aureus, the expression levels of proinflammatory cytokine genes, including IL-4, IL-13, and thymic stromal lymphopoietin (TSLP), are stimulated and the Th1/Th2 immune response is induced.5052 Colonization with S. aureus could impair the suppressive activity of Treg cells.5354 In addition, the presence of S. aureus in the skin is related to a high IgE response.53 These findings show that S. aureus flares in the skin exacerbate the skin disease by further promoting inflammation. In addition, in patients with AD, marked reductions in skin microbial diversity occur during disease flares and effective treatment restores diversity to the skin bacterial community. Flares are characterized by increased abundance of pathogenic S. aureus.5556
Most studies of the skin microbiome in AD have focused on S. aureus, and a few studies have shown that Staphylococcus epidermidis can inhibit the growth of S. aureus; additionally, colonization with S. epidermidis and S. cohnii reduced the development of AD in children at age 1.5657
However, in recent studies, skin immunity was differentially affected by the time of S. aureus appearance. In a mouse model, cutaneous exposure to commensal bacteria early in life induced tolerance to S. aureus.58 Another birth cohort study showed a difference in staphylococcal colonization, namely fewer Staphylococcus species, at a site of AD predilection that was later affected by AD.47 These observations suggesting that the timing of exposure to commensal bacteria influences the development of tolerance prompted us to explore how neonatal and adult skin differs with respect to the resident immune cell populations. Neonatal skin Treg cells are more activated and abundant than their adult counterparts, constitute the majority of T cells in the skin during this key developmental window, and are unique to the skin versus another key barrier site, the gut.59 As long as exposure to various food antigens at younger ages causes tolerance, exposure to S. aureus at an early age on the skin may help induce tolerance because gut exposure to diverse food antigens at a younger age causes tolerance.
To confirm the characterization of community function and genomes, metagenome shotgun analysis of human skin was needed. A skin metagenome shotgun study showed an interaction between biogeography and individuality shape function in human skin60 as well as a cross-modulated interaction between skin microbial communities, the skin surface microenvironment, and the immune system in AD flares.61 AD-associated microbiomes can increase the risk of flares by influencing the skin surface microenvironment and through an interaction with the host immune system. Future functional studies of the skin microbiome at the metatranscriptomic, metaproteomic, and metabolic levels are vital for our understanding of AD mechanisms and may identify future therapeutic targets of the microbiome in AD and skin health maintenance.
Go to :
The effects of skin immunity on systemic and intestinal immunity
Studies of the effects of skin immunity on the gut and systemic immunity have been performed in the food allergy field, but not for AD. Food allergy and AD are closely related, and, epidemiologic studies have shown that AD-associated cutaneous inflammation is a significant risk factor for the development of food allergy.6263 Exposure to food allergens via the cutaneous route and its extremely efficient antigen-presenting cells, before exposure via the oral route, bypasses oral tolerance. Thus, when the gut does get exposed to food allergens, this previous sensitization by the cutaneous route leads to symptoms associated with food allergy.64 The immunologic mechanisms by which antigen sensitization in the skin can predispose the gut to allergic inflammation are incompletely understood, but a recent animal study showed that epicutaneous sensitization on a disrupted skin barrier is associated with accumulation of TSLP-elicited basophils and DCs, which are necessary and sufficient to promote antigen-induced food allergies.65 Injured epithelial cells may also release IL-33 to activate ST2-expressing skin DCs.66 TSLP-activated DCs migrate to draining lymph nodes, which in turn induce naïve CD4+ T cells to differentiate into CD4+ Th2 cells and maintain CD4+ Th2 effector/memory pools.67 Gut IL-25 propagates the allergic immune response by enhancing collaborative interactions between resident type 2 innate lymphoid cells and CD4+ Th2 cells expanded by ingested antigens in the gut.68 IL-4 signaling provided by CD4+ Th2 cells induces emigrated mast cell progenitors to become multifunctional IL-9-producing mucosal mast cells, which then expand greatly after repeated food antigen exposure.69
On the other hand, cutaneous exposure, such as epicutaneous immunotherapy (EPIT), may be used to treat food allergy. EPIT is a novel experimental method that attempts to induce tolerance in allergic patients by delivering low concentrations of food allergens through healthy skin. In milk-sensitized mice, EPIT induced milk-specific Treg cells that subsequently promoted tolerance to cutaneously administered peanuts and house dust mites.70 The results are similar to those of subcutaneous and sublingual routes of inhalant allergen exposure, where allergen tolerance is attributed to the induction of Treg cells and IL-10.71
How the same skin exposure can either induce intestinal sensitization or cause immune tolerance has not been elucidated. However, differences in exposure time and the presence of microorganisms in the skin or gut are likely to be involved and need to be examined further.
Go to :
CONCLUSIONS AND PERSPECTIVES
Here, we explored the current evidence for the existence of a translationally relevant gut-skin axis. The roles played by the gut and skin microbiome have recently been revisited and abundant evidence suggests that host-microbiome interactions can actually determine the status of health and AD. It is unclear whether changes in the microbiome at one organ site affect distal organs or different organ sites and whether these systemic effects might be specific for certain tissues or organs, and the mechanisms involved. To date, no broad study investigating these systemic widespread effects has been performed.
S. aureus species of the skin microbiome play key roles in the development and establishment of AD, but the proportional differences among other skin microbes are implicated in AD development. Therefore, extensive research is required to identify the interactions between the skin microbiomes or between the gut and skin microbiomes and how the mechanisms involved influence the development of AD dysbiosis. Moreover, additional investigations are needed to confirm that AD dysbiosis through the skin microbiome affects the skin response and systemic immune response in AD. It is also unclear whether S. aureus species or other microbes may have some function in the gut.
The compositional and proportional differences in the gut microbiome are associated with the development of AD via an immunomodulatory effect of the gut microbiome. The gut microbiome may contribute to the development, persistence, and severity of AD via immunologic, metabolic, and neuroendocrine pathways. To identify the in-depth role of the gut or skin microbiome in the development of AD, studies should investigate both the gut and skin microbiome and the diverse pathways involved in cross-talk between the gut or skin microbiome and the immune system.
Metagenomic analyses in humans and animals clearly demonstrated that AD is associated with the dysbiosis of the gut and skin microbiome. However, causal relationships between microbiome and AD have rarely been elucidated. In this regard, microbiome, metatranscriptomic, metagenomic, and metabolomic analyses can be promising tools for understanding the etiology and increasing prevalence of AD in recent decades. Furthermore, identification of novel microbial genes and molecular pathways capable of modulating mucosal Th responses might be immensely helpful in achieving in-depth understanding of the origin of AD.
Go to :
 XML Download
XML Download