

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The rapidly rising prevalence of food allergy worldwide has now been termed the “second wave of the allergy epidemic.”1 Up to 3% of children in the US2 and 9.5% of Australian infants aged 11–15 months3 are now allergic to peanuts and eggs, respectively. Recent high-profile randomized controlled trials in the US, UK, Australia, and Japan have examined the role of early introduction of allergenic food, such as peanuts and eggs for the prevention of peanut and egg allergies respectively.45 New infant feeding guidelines were also released in the US, UK, and Australia, recommending early introduction of peanuts in high-risk infants for the prevention of peanut allergy.678
However, the epidemiology of food allergy, its risk factors, diagnostic techniques, and efficacy of preventive measures are remarkably variable across different populations throughout the world, particularly between Asian and Western societies. Gene-environment interactions may modulate the pathogenesis of allergic disorders, including food allergy and potentially influence each population's response to the same interventions. This review explores geographical variations in food allergy epidemiology and risk factors in different populations, and discusses their implications in clinical practice.
Food allergy patterns across various populations
The epidemiology of food allergy paints a contrasting picture across countries worldwide (Figure). Studies on childhood food allergy found that the overall prevalence of oral challenge-proven food allergy in children below 5 years of age was only 1% in Thailand,9 but as high as 5.3% in Korean infants10 and 10% in Australian pre-schoolers.3
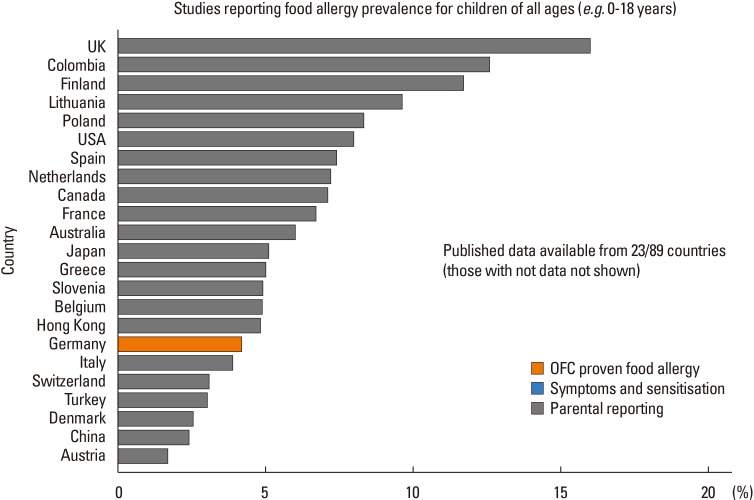 | FigureStudies are categorized according to level of evidence; OFC proven food allergy (orange bars); or questionnaires/parental reporting (grey bars). Figure from Prescott et al. A global survey of changing patterns of food allergy burden in children. World Allergy Organization Journal. 2013 Dec 4;6(1):21. doi: 10.1186/1939-4551-6-21. BioMed Central Ltd.
|
While milk and eggs are the most common allergens in early childhood in the UK, US, Australia, and many parts of Europe and Asia, distinct differences in prevalence are observed even between countries located within the same continent. The EuroPrevall birth cohort, which recruited infants from 9 European centers with different climatic and cultural backgrounds, found that the incidence of challenge-proven cow's milk allergy was lower in southwestern European countries, such as Greece (0%) and Italy (0.3%), but was highest in the UK (1.24%).11 The prevalence of egg allergy was also variable—the lowest incidence was again reported in Greece (0.07%) and the highest in the UK (2.18%).12
The common food allergens in Asia are also different from those seen in Western countries. Peanut allergy predominates in children less than 5 years of age in the UK, US, and Australia,13 but is very uncommon in the rest of Asia apart from Japan.14 Wheat is becoming one of the most common food allergens in childhood, followed by egg and milk, in Thailand, Japan, and Korea.15 A multicenter study in Korea found that walnuts were the most common cause of physician-diagnosed food allergy in preschoolers and school-age children, whereas buckwheat and wheat allergy were most common in adolescents.16 Shellfish is also a leading cause of food allergy in adolescents and adults in other parts of Asia.17
Ethnic-dependent variability for food allergy risk is evident even within the same country. In South Africa, black African (Xhosa) children had a significantly lower prevalence of peanut allergy compared to children of mixed race origin (Caucasian and Black).18 African-American and Hispanic children living in the US had significantly higher odds of physician-diagnosed food allergy and eczema compared to white children.19
Migrant studies also suggest a strong environmental influence on food allergy risk. Du Toit et al.20 found that Jewish children living in the UK who avoided peanuts in the first year of life had a 10-fold higher risk of peanut allergy compared to Jewish children living in Israel, in whom peanuts were introduced early in infancy.20 The HealthNuts study recruited 5,276 infants in Melbourne, Australia, screened them with skin prick tests, and subsequently performed oral challenges for gold-standard confirmation of food allergy. The investigators observed a significantly increased risk of peanut allergy in infants with parents who were born in East Asia (7.7%, OR 3.4, 95% CI 2.2–5.1) compared to those whose parents were born in the UK or Europe (2.3%).21 Panjari et al.22 likewise observed that children born in Australia to Asian mothers had a higher risk of parental-reported peanut allergy (OR 2.9, 95% CI 2.17–3.89) compared to Australian-born white children. Birth in Asia and postnatal migration to Australia was in fact protective against nut allergy in this cohort. A population-based questionnaire survey on peanut and tree nut allergy in Singapore and the Philippines found that both ethnicity and place of birth were important risk factors for peanut and tree nut allergy.23 Children of either Asian or white ancestry who were born in Western countries had an increased risk of peanut allergy compared to those who were born in Asia (OR 3.47, 95% CI 1.35–8.93). Ethnic Asian children born in Western countries had a 5-fold higher risk of tree nut allergy compared to Asia-born Asian children. In contrast, white children born in Asia had a significantly lower risk of nut allergy than expatriate white children (OR 0.24, 95% CI 0.07–0.74).
Migration and duration of residence in the host country also appear to impact upon AD risk, which might explain the higher prevalence of food allergy in Australian immigrants. Infants with East Asian ethnicity who had migrated to Australia were found to have a significantly higher risk of eczema compared to infants of Australian-born parents.24 The proportion of East Asian infants with eczema was also higher in those whose parents had migrated to Australia more than 5 years before study recruitment, compared to those who had migrated more recently.
Aeroallergen-food cross-reactivity syndromes
Geographical variations in the distribution of aeroallergens also influence food allergy patterns where cross-reactivity between aeroallergen and food allergen components exist. These aeroallergen-food cross-reactivity syndromes occur when IgE sensitization to aeroallergens first develops through the respiratory route and subsequently causes clinical allergy to food allergens which share molecular homology with the implicated aeroallergens. Clinical symptoms may range from the oral-allergy syndrome to full-blown anaphylaxis.
IgE sensitization patterns to aeroallergens and their respective cross-reactive food allergens have been shown to be variable according to geographical location. A study of 115 peanut-allergic children from 3 countries—Spain, the US, and Sweden found that the majority of American patients were sensitized to Ara h 1, 2, and 3 (seed storage proteins) and had an earlier onset of peanut allergy and more severe clinical symptoms. Rates of sensitization to birch and grass pollens were very low in this group.25 In contrast, Spanish patients were more often sensitized to the lipid transfer protein (LTP) Ara h 9 and developed peanut allergy later than American subjects, often only after initial sensitization to cross-reactive plant-derived foods. Swedish children had the highest sensitization rate to Ara h 8 and its homologue—the birch pollen allergen Bet v 1. These differences might be attributed to variable environmental pollen exposures as well as culinary practices unique to each population.
The EuroPrevall study evaluated 731 subjects across 12 European cities with hazelnut allergies and found that hazelnut sensitization was mainly birch pollen-driven (Cor a 1) in subjects from Central and Northeastern Europe.26 However, Cor a 8 (LTP) sensitization was predominant in the Mediterranean cities and was associated with concomitant sensitization to walnut, peach, apple, and other tree pollen. Sensitization to hazelnut seed storage proteins (Cor a 9, Cor a 11, and Cor a 14)—primary sensitization—was observed in less than 10% and mostly in children who were also sensitized to nuts, seeds, and legumes.
The EuroPrevall group also found that kiwifruit allergic patients in western, central, and eastern Europe were mainly sensitized to Act d 8 (pathogenesis-related class 10 protein: PR-10) which is related to birch pollinosis, whereas those in southern Europe were sensitized to Act d 9 (profilin) and Act d 10 (non-specific LTP). Patients from Iceland, were however, more likely to be sensitized to Act d 1 (LTP) and also manifested with more severe clinical symptoms.27
The prevalence of shellfish allergy ranges from 0.1% to 5.5% worldwide, predominantly in adolescents and adults, and appears to be more common in Asian than Western populations.13 Environmental factors, such as house dust mite (HDM) exposure, could contribute to differences in the prevalence of shellfish allergy. The HDM-crustacean syndrome is an emerging phenomenon in which primary sensitization to dust mite allergens induces allergic reactions to shellfish through cross-reactive tropomyosin allergens. Tropomyosin is an integral component of actin filaments, which regulate muscle contraction in invertebrates, such as dust mites, shellfish, mollusks, and other insects. Tropomyosin Der p 10 is the most common allergen component shared by these organisms and has been implicated as the main sensitizer in mite-crustacean cross-reactivity.28
Shellfish sensitization highly correlates with HDM sensitization in warm humid climates and urban environments where HDMs are ubiquitous. Inhibition assays in shrimp allergic Spanish patients found that HDMs were primary sensitizers in subjects living in humid climates, whereas those living in dry climates were sensitized only to shrimp allergens.29 Another study in Singapore observed that shrimp-allergic subjects were also all dust mite-sensitized. In fact, a subgroup of patients were only sensitized to HDM without any evidence of sensitization to shrimp itself, suggesting that HDMs were likely primary sensitizers in this group.30 Jewish subjects who adhered to strict lifetime dietary avoidance of shellfish were also found to be sensitized to shrimp through cross-reactivity to HDM tropomyosin allergens.31 It is thus likely that the prevalence of shellfish allergy correlates closely with environmental exposure to HDM, but more prospective studies are needed to investigate the temporal relationship between HDM exposure, sensitization, and the development of shellfish allergy.
Genetics and epigenetics
Familial aggregation, heritability estimates, and twin studies indicate that a genetic link to peanut allergy exists. Candidate gene studies have identified specific HLA haplotypes to be associated with peanut allergy. Howell et al.32 found that HLA-DRB1* 08 and DQB1*04 alleles were increased in frequency among peanut-allergic subjects compared to healthy unrelated controls. Technological advances subsequently paved the way for genome-wide association studies (GWAS) which broadened the search for specific genetic heritability markers for peanut allergy. The Chicago Food Allergy Study conducted the first GWAS study on food allergy, which identified peanut allergy-specific loci in the HLA-DR and -DQ gene region on chromosome 6p21.32 which were tagged to the 2 single nucleotide polymorphisms (SNPs) rs7192 and rs9275596 in subjects of European ancestry.33 These SNPs were associated with differential DNA methylation levels at multiple CpG sites, which partially mediated the association between SNPs and peanut allergy. These associations were, however, not present in subjects of non-European ancestry from the same cohort, though this could potentially be due to underpowering stemming from the small sample size. It is not known whether this association exists in other ethnic groups as no GWAS studies on food allergy have thus far been performed outside of the white population.
Filaggrin (FLG) loss-of-function mutations have been shown to be associated with atopic dermatitis (AD) and other allergic disorders, such as peanut allergy. The filaggrin gene encodes an essential epidermal protein expressed in the outer layers of the epidermis and plays an essential role in maintaining skin barrier integrity. The dual-barrier hypothesis proposes that a disrupted skin barrier in early life, such as in AD and filaggrin loss-of-function mutations, predisposes to allergic sensitization to food and environmental allergens, while early oral exposure to food allergens promotes tolerance.
There are considerable differences in the carrier frequency and types of FLG mutations in AD patients from different ethnic groups. Up to 50% of European patients with AD had FLG mutations, mainly involving 2 mutations (R501X and 2282del4).34 In contrast, only around 20% of Asian AD subjects carry FLG mutations, but these covered a wider spectrum of genotypes distinct from the European population.34 Chen et al.34 observed 14 novel FLG-null mutations in Singaporean Chinese patients with AD and ichthyosis vulgaris, which were not found in European cohorts. Eight unique FLG-null mutations predominated in these patients compared to only 2 in the Irish cohort. Studies in Asia showed that the prevalent FLG mutations in each Asian country were also highly variable and only the 3321delA mutation was common to all.3435
There are few studies examining the direct association of FLG mutations with food allergy. FLG loss-of-function mutations were found to be associated with peanut allergy in English, Dutch, and Irish patients, even after controlling for AD.36 Brough et al.37 also described a dose-response relationship between environmental peanut exposure in the first year of life and subsequent peanut sensitization in children with FLG null mutations. The Consortium for Food Allergy Research (CoFAR) study in the US, however, reported a low prevalence of FLG mutations in peanut-allergic/sensitized children, and FLG mutations were not associated with peanut allergy outcomes.38 Nomura et al.39 identified an association between a common FLG variant (rs 1933064) and food sensitization in Japanese infants, but there was no association between combined FLG null mutations and food sensitization or AD itself. The Australian HealthNuts study likewise found that FLG mutations did not increase the risk of food allergy over and above that of food sensitization,40 suggesting that while FLG mutations might increase the risk of sensitization through an impaired skin barrier, in some populations other factors are required for progression to clinical food allergy.
The Asian AD phenotype comprises well-demarcated, erythematous plaque-like skin lesions, whereas European-American subjects display ill-defined, macular erythematous skin lesions.41 Both Th2 and Th17 cytokines drive Asian AD, while AD in European-Americans is mediated mainly by Th2 cytokines alone. The relationship between genetics, epithelial immune responses, and clinical phenotypes between populations are, however, yet to be fully elucidated.
Other genetic markers are now being recognized as potential predictors of food allergy. The HealthNuts study recently identified associations between a polymorphism in IL-13 (rs1295686)42 and SPINK5 genetic variants43 with challenge-proven food allergy in Australian infants. Associations in other populations have not yet been established.
Epigenetic modifications due to environmental influences may also be key mediators of food allergy risk, which may vary between populations. In vivo studies have demonstrated that the inter-generational inheritance of epigenetic modifications is possible which supports the hypothesis that environmental influences may exert selection pressures conferring differential allergy-related modifications to the epigenome, which could then be passed down to offspring pre- or perinatally. This mechanism might partially explain the effects of environmental selection pressures due to migration on food allergy risk observed in different ethnic groups.
Meteorological factors and vitamin D status
Ecological studies have identified several climatic factors, such as latitude, sunlight exposure, and season of birth, correlating with Vitamin D status, as well as ambient humidity, as potential geographic determinants that may modulate food allergy risk in infancy. An early study by Camargo et al.44 first reported a latitude-dependent gradient for adrenaline autoinjector prescriptions, used as a surrogate marker of anaphylaxis prevalence, in the US, with a 4-fold higher rate of prescriptions in the northern states compared to the south. This observation was also consistent with that in Australia, where higher rates of EpiPen® prescriptions and hospital admissions for anaphylaxis were found in southern Australia compared to the northern states which are closer to the equator.45 As ultraviolet radiation exposure, which induces vitamin D synthesis, decreases with distance from the equator, it was hypothesized that low vitamin D status could be an important risk factor for food allergy development.
Ambient UV exposure is lower during fall and winter. It would thus likely be reflected in lower vitamin D levels in infants born during these seasons. Season of birth studies that examined this hypothesis in further depth, however, showed conflicting results. Keet et al.46 reported increased rates of food allergy or food sensitization in infants born in fall or winter seasons in the US. Thysen et al.47 further found that infants born in fall and winter had higher levels of activated and regulatory T cells in their cord blood compared to infants born in summer. These studies were, however, limited by the lack of data on exogenous vitamin D intake, maternal vitamin D intake and measurement of vitamin D levels in the infants.
In contrast, Kusunoki et al.48 noted that the initial observation of an association between autumn/winter births and food allergy in Japanese infants was confounded by the existence of AD before 6 months of life, postulating that season of birth was more likely a risk factor for AD than food allergy per se.
Apparent discrepancies between studies on vitamin D status and food allergy associations49 may be explained by ethnic-dependent genetic polymorphisms in vitamin D-binding protein, which evoke differential effects on vitamin D bioavailability. Keet et al.46 found that the season of birth effect was significant for food allergy only in Caucasian infants, but not in non-Caucasian infants. African-American subjects in the ‘Healthy Aging in Neighborhoods of Diversity across the Life Span’ cohort had lower total 25-hydroxyvitamin D levels than white American subjects, but levels of bioavailable Vitamin D were comparable between both groups. This was found to be due to genetic polymorphisms in the vitamin D-binding protein gene (rs7041 and rs4588) which resulted in reduced Vitamin D-binding protein levels in African-Americans, translating to a relatively higher amount of circulating bioavailable vitamin D in these subjects.50 The HealthNuts cohort first reported an association between vitamin D insufficiency (≤50 nmol/L) and peanut and/or egg allergy in infants of Australian-born parents, but not of parents born overseas, in particular East Asia.51 They subsequently found that these same genetic polymorphisms modulated the above-mentioned associations between vitamin D insufficiency and food allergy risk in their cohort. Infants with parents born in Asia were more likely to have genotypes predictive of low vitamin D-binding protein levels, which could have negated the adverse effects of vitamin D insufficiency on food allergy risk in this ethnic group.52
Dietary patterns
Cultural dietary practices are widely variable between populations. Culinary methods might partially explain the differences in food allergy prevalence between East and West. Roasted peanuts, which are common in Western diets, confer higher allergenicity compared to boiling or frying,53 which is practiced more often in Asian cuisine.53 Treatment of peanut with food additives, such as vinegar in Korea, has also been shown to reduce peanut allergenicity.54
Changes in infant feeding practices over the past 2 decades have been postulated to be partly responsible for the exponential rise in food allergy rates. In the year 2000, the World Health Organization (WHO) recommended that complementary foods be introduced no earlier than 6 months of age.55 The American Academy of Pediatrics (AAP) further recommended maternal restriction of peanuts and tree nuts, and the elimination of cow's milk, eggs, and fish during lactation to reduce the risk of food allergy in their offspring. It also recommended that solid foods not be introduced into the diet of high-risk infants until 6 months of age and dairy products delayed until 1 year, eggs until 2 years, and peanuts, nuts, and fish until 3 years of age.56 However, as evidence for delayed allergenic food introduction for food allergy prevention was lacking, the AAP & the National Institute of Allergy and Infectious Diseases (NIAID) subsequently replaced these guidelines with new recommendations that solid foods not be delayed beyond 4–6 months of age and that the introduction of allergenic foods into infants' diets should no longer be delayed.57
Since then, several observational studies and randomized controlled trials have examined the role of early allergenic food introduction into the diets of normal and high-risk infants, but results were conflicting depending on the individual study populations and choice of food allergen introduced (Table 1). Katz et al.58 found that infants in Israel who were exposed to cow's milk in the first 14 days of life were less likely to become cow's milk-allergic compared to those who had their first exposure beyond the first 14 days. Israeli children who consumed peanuts in high quantities from early life had 10 times lower risk of peanut allergy compared to Jewish children living in the UK who tended to avoid peanuts.20 The Australian HealthNuts cohort study also found that early introduction of cooked egg into infants' diets between the age of 4 to 6 months reduced the risk of egg allergy, while delayed introduction after 12 months of age was associated with higher rates of egg allergy (adjusted odds ratio 3.4, 95% CI 1.8 to 6.5).59 In contrast, the Growing Up in Singapore Towards healthy Outcomes (GUSTO) cohort study, showed that the prevalence of peanut allergy was very low (0.1%–0.3%) in children aged 12–48 months despite delayed introduction of peanut beyond 10 months of age in 88% of the study population.60
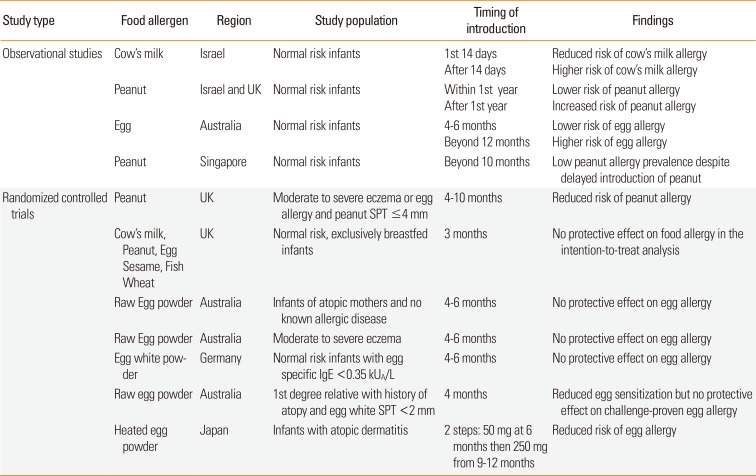
Table 1
Timing of allergenic food introduction and food allergy
![]()
The Learning Early About Peanut Allergy (LEAP) trial enrolled 640 high-risk infants with severe AD and/or egg allergy in the UK between the ages of 4 and 11 months, who had peanut skin prick test (SPT) responses of <4 mm. They were randomized to either consume 6 grams of peanut protein at least 3 times a week or to completely avoid peanut products for the first 5 years of life. In the intention-to-treat analysis, 17.2% of children in the peanut avoidance group developed challenge-proven peanut allergy by age 5 years, compared to only 3.2% in the peanut consumption group, translating to a risk reduction of 86.1%.61
The Prevention of Egg allergy with Tiny amount InTake (PETIT) study was a randomized double-blind placebo-controlled trial which investigated the effects of early introduction of heated egg powder in high-risk Japanese infants with AD on the development of egg allergy.62 The intervention group received 50 mg heated whole egg powder between 6–9 months of age, which was then increased to 250 mg between 9 and12 months of age. The authors showed a significantly reduced risk of hen's egg allergy through open food challenge at 12 months of age (8%, 5/60) compared to the placebo group (38%, 23/61), with a risk difference of 29.4% (95% CI 15.3–43.4) and a number needed to treat of 3.40 (2.30–6.52).
The Enquiring About Tolerance (EAT) study randomized normal risk, fully breastfed 3 months old infants in the UK to the early introduction group: regular consumption of 6 allergenic foods (peanut, cooked egg, cow's milk, sesame, white fish, and wheat) or to the standard introduction group: to continue exclusive breast-feeding to around 6 months of age before introducing allergenic foods. However, the intention-to-treat analysis found no significant protection of early allergenic food introduction against food allergy development.63 Several randomized trials on early egg introduction in Australia and Germany also failed to demonstrate a protective effect against egg allergy development in both high-and normal-risk infants.5 There was also considerable heterogeneity between these studies, namely the egg preparation methods which varied from raw to heated egg, as well as whole versus egg white powder, which could have explained the lack of significant findings in some of these studies.
Implications for clinical practice
Diagnostics
Component resolved diagnostic methods are increasingly being used in the diagnosis and management of food allergies. However, the utility of various allergen components in predicting clinical reactivity differs significantly between populations. Studies have demonstrated the superior performance of sIgE to the peanut component Ara h 2, a seed storage protein, in predicting peanut allergy, suggesting that the test might potentially obviate the need for high-risk diagnostic food challenges. However, the sensitivity and specificity of this test range from 60%–100% and 60%–96%, respectively, using a cutoff of 0.35 kU/L, depending on the population in which it is used.64 As discussed earlier, the pattern of sensitization to peanut components varies between peanut-allergic populations.25 In countries like the Mediterranean region, sensitization to other peanut components, such as Ara h 9, appears to be a better predictor of clinical reactivity than Ara h 2.65 Gray et al.18 also found that Ara h 2 had a significantly lower predictive value for peanut allergy in native Africans (53%) compared to those of mixed race (93%).
Performance of component-resolved diagnostics for hazelnut allergy also differs between populations. Challenge-proven hazelnut allergic subjects in Denmark and Switzerland most commonly had sIgE to recombinant Cor a 1, while those in Spain mainly had sIgE to recombinant Cor a 8.66 A Dutch study found that sensitization to Cor a 9 and Cor a 14 instead was highly specific for challenge-proven hazelnut allergy and was also a marker of increased symptom severity.67
Studies of 95% positive predictive values (PPV) for skin prick tests and specific serum IgE for predicting positive oral food challenges show substantial variability between populations due to differences, such as age, ethnicity, atopic risk as well as assay techniques.
US children aged 3 months to 14 years referred for suspected food hypersensitivity were found to have 95% positive predictive values of 7 kUA/L for egg and 15 kUA/L for milk for failing oral challenges.68 However, threshold values are quite different in other populations: 1.5 kUA/L (95% PPV for eggs) in Danish children younger than 2 years and 1.3 kUA/L in those older than 2 years, referred for suspected egg allergy69; while in Japanese children, 95% PPVs for failing egg challenges were 13 kUA/L, 23 kUA/L, and 30 kUA/L in children less than 1 year, 1 year of age, and above 2 years, respectively. The 95% PPVs for milk were 5.8 kUA/L, 38.6 kUA/L, and 57.3 kUA/L, respectively.70
Predictive thresholds for skin prick tests are also equally variable and some studies were performed in populations with relatively higher food allergy prevalences, which result in high pretest probabilities and higher corresponding PPVs, which may not be applicable to other populations with a lower prevalence of food allergy. In the absence of universally validated threshold values for these tests, diagnostic approaches and clinical decision-making processes have to be individualized to each population.
Food allergy prevention
A joint consensus communication was released by several Allergy organizations worldwide in 2015, recommending introduction of peanut-containing products into the diets of “high-risk” infants early in life (between 4 and 11 months of age) in countries where peanut allergy is prevalent because delaying the introduction of peanut can be associated with an increased risk of peanut allergy.”8 US addendum guidelines released in January 2017 further provided 3 risk-stratified recommendations for peanut introduction in infants in the US6:
Infants with severe eczema, egg allergy, or both - to strongly consider evaluation by sIgE measurement and/or SPT and, if necessary, an OFC. Based on test results, introduce peanut-containing foods.
Infants with mild to moderate eczema - to introduce peanut-containing foods around 6 months of age.
Infants without eczema or food allergy - to introduce peanut-containing foods in an age-appropriate manner and in accordance with family preferences and cultural practices.
However, given the aforementioned differences in food allergy prevalence, allergenic food introduction practices and food allergy associations in nonwhite populations, it is debatable whether these guidelines on peanut allergy should be equally applicable to other ethnic groups, as well as in countries where the burden of peanut allergy is considerably lower than in the trial populations. There is also currently no consistent evidence to show that early introduction of any other food allergen, such as milk, soy, wheat, or shellfish is protective against food allergy development.
Conclusions and future directions
The variability of food allergy characteristics between populations can be attributed to a complex interplay of genetic, as well as environmental influences, rather than any single factor alone, as summarized in Table 2. Interventions aimed at preventing food allergy development and managing established food allergy must thus be individualized to each population, based on each population's unique characteristics.

Table 2
Factors relating to differences in food allergy around the world
![]()
More research is needed to determine the indications for early allergenic food introduction and the choice of allergens to be introduced early across different populations. Moreover, primary preventive strategies may need to be prioritized differently between populations. For example, prevention of HDM sensitization might be more important for the prevention of shellfish allergy in at-risk populations where shellfish allergy and HDM sensitization rates are high, because of the likelihood of mite-crustacean cross-reactivity. In populations where there is little evidence to support early dietary interventions, efforts might instead be directed towards the primary prevention of eczema, such as through preemptive application of emollients from birth in high-risk infants and proactive management of established AD for secondary prevention of sensitization and progression to clinical food allergy. The risk of changing existing infant feeding practices in a population where the specific food allergy prevalence is already low may not be justified, as it may unwittingly alter other lifestyle practices or unknown environmental factors that may conversely result in the opposite undesired effect.
Studies on the utility of component-resolved diagnostics in other food allergens, beyond peanut, need to be validated in more diverse populations. Future research should also include studies on early introduction of allergenic foods, beyond peanut, such as milk, wheat, and shellfish into the diets of high-risk infants, within the populations where these allergies are highly prevalent. There is also a need to standardize study methodologies, such as the type and preparation of the allergenic food—for example, raw versus heated egg powder, so that findings can be compared between populations. Genetic markers for food allergy susceptibility and response to dietary interventions also need to be identified. More research is also required to elucidate the role of HDM sensitization in progression towards shellfish allergy and the efficacy of preventive interventions in environments where HDM prevalence is high.
Go to : 
 XML Download
XML Download