

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Adenine, a purine base, is a primordial molecule that is a component of essential biomolecules such as nucleic acids and adenine nucleotides and is a modulator of various physiological functions [123]. Proliferating cells make 5-methylthioadenosine (MTA) during polyamine synthesis [4]. MTA-phosphorylase yield 5-methylthioribose-1-phosphate and adenine and contributes 85–97% of total adenine synthesis in replicating cells [5]. Adenine is cell-permeable and convertible to adenosine monophosphate (AMP) via adenine salvage pathway. AMP and ATP pools are replenished during the salvage thereby contributing to its cellular and physiological activities [4]. AMP-activated protein kinase (AMPK) is activated by phosphorylation at Thr-172 residue by Liver Kinase B (LKB). AMP activates AMPK by inhibiting dephosphorylation of phosphorylated AMPK as well as acts as an allosteric activator of AMPK [6]. Also, G protein-coupled receptors (GPCR) specific to adenine have been cloned and characterized in mouse brain [78] and pig kidney [9]. Therefore, various biological functions of adenine have been reported. It increases survival of rat Purkinje cells [3] and erythrocytes in whole blood storage [10] and modulates contraction of rat thoracic aorta [11]. It also regulates immune responses such as human lymphoblast growth [212], mitogen-stimulated transformation, cytoplasmic immunoglobulin production, natural killer activity of human mononuclear leukocytes [113] and IgE-mediated mast cell activations [14]. Adenine inhibits the TNF-α signaling in intestinal epithelial cells and HUVECs through PKA activation and AMPK activation respectively [15].
Lipopolysaccharide (LPS), a cell wall component of Gram-negative bacteria, is a well-known activator of toll like receptor 4 (TLR4) which can evoke uncontrolled inflammatory responses, organ failure and death [1617]. LPS-induced production of inflammatory cytokines in the circulation system causes development of systemic inflammatory response syndrome (SIRS) and sepsis [1819]. Innate immune cells, such as macrophages and mast cells express Toll-like receptor 4 (TLR4) which recognize LPS. Upon forming the complex with CD14, LPS trigger serial signal transduction events which lead to the activation of nuclear factor-κB (NF-κB), mitogen-activated protein kinase (MAPK) and cytosolic phospholipase A2 (cPLA2). Activation of these proteins leads to the production of inflammatory cytokines such as necrosis factor-α (TNF-α) and interleukin-6 (IL-6) and eicosanoids such as prostaglandin E2 (PGE2) and leukotriene B4 (LTB4) thereby initiating the inflammatory reactions [2021]. JUN Nterminal kinase (JNK) 1/2, p38 MAPK and extracellular-signal-regulated kinases (ERK) 1/2 are components of MAPK pathway which regulates NF-κB and activator protein-1 and consequently synthesis of inflammatory cytokines and mediators in LPS-stimulated conditions [22]. Phosphoinositide 3-kinase (PI3K)/Akt signaling, along with NF-κB and MAPK is also important in regulation of cytokine production because Akt activation leads to NF-κB mediated cytokine production [2324]. The pro-inflammatory cytokines further lead to activation of immune cells and worsen the inflammation. Macrophages [2526] and mast cells [2728] react against LPS via TLR4 and then secrete pro-inflammatory cytokines. Hence inhibition of these signaling pathways leads to reduction in pro-inflammatory mediators. However, adenine effect on LPS-induced inflammatory reactions is not studied.
In this study we explored adenine roles in LPS-induced inflammatory reactions. Adenine inhibited the LPS-induced production of pro-inflammatory cytokines and lipid mediators in RAW264.7 cells and bone marrow derived mast cells (BMMCs). Adenine inhibited activation of NF-κB, MAPK and Akt signaling and enhanced cellular AMP level without activation of AMPK. Also, adenine attenuated LPS-induced production of TNF-α and IL-6 by peritoneal cells in mice.
METHODS
Materials
The following materials were purchased from the indicated commercial sources: adenine, hypoxanthine, lipopolysaccharide (LPS), methyl thiazolyl tetrazolium (MTT), Dimethyl sulfoxide (DMSO), 2-mercaptoethanol, anti-β-actin antibody from Sigma-Aldrich (St Louis, MO, USA); fetal bovine serum (FBS) from Gibco/Life Technology (Grand Island, NY, USA); Dulbecco's Modified Eagle Medium (DMEM) from Welgene (South Korea); recombinant IL-3 from Peprotech (Rocky Hill, NJ, USA); RPMI 1640 medium from Lonza (Walkersville, MD, USA); anti-NF-κB p65, anti-phospho-NF-κB p65 (Ser536), anti-phospho-IκB (Ser32), anti-Akt, anti-phospho-Akt (Ser473 and Thr308), anti-ERK and anti-phospho-ERK (Thr202/Tyr204), anti-phospho-p38 MAPK (Thr180/ Tyr182) and anti-p38 MAPK antibodies from Cell Signaling Technology (Beverly, MA, USA); anti-phospho-JNK (Thr183/Tyr185) and anti-JNK antibodies from Invitrogen (Rockford, IL, USA); Thermo Scientific NE-PER nuclear and cytoplasmic extraction reagents from Pierce Biotechnology (Rockford, IL, USA); ELISA sets for mouse TNF-α, IL-6 and IL-13 cytokine from BD Biosciences (San Diego, CA, USA); Prostaglandin E2 Assay kit from R&D system (Minneapolis, MN, USA); Leukotriene B4 assay kit from Enzo life sciences (Farmingdale, NY, USA); AMP-Glo assay kit from Promega (Madison, WI, USA); RIPA lysis buffer from ELPIS Biotech (South Korea); ECL chemiluminescence kit from Millipore (Billerica, MA,USA); RAW264.7 cell line from American Type Culture Collection.
Cell culture
RAW264.7 cells were cultured in DMEM supplemented with 10% FBS and standard concentration of penicillin/streptomycin antibiotics. Cultures were maintained in a standard condition in 5% CO2 incubator at 37℃. For all experiments, required numbers of cell were incubated overnight in appropriate plates. Then various doses of adenine were treated for 30 min in a complete growth medium before stimulating cells with LPS (100 ng/ml) for indicated time and cells were harvested according to required experimental purposes.
ELISA to measure cytokines and eicosanoids
RAW264.7 cells (1×105 cells per ml per well) were seeded in 12-well cluster plates and incubated overnight. Then cells were stimulated with LPS after pre-incubation with adenine or hypoxanthine for 30 min. Levels of cytokines and eicosanoids in the supernatants were measured using the ELISA kit according to the manufacturer's protocol.
Cell viability assay
Cell viability was determined using thiazolyl blue tetrazolium bromide (MTT). Cells seeded into 96-well plates were incubated overnight at 37℃ and then treated with various doses of adenine for 6 h. The cells were then incubated with MTT (0.5 µg/ml) for 2 h and the formazan produced was dissolved in DMSO. Absorbance was measured at 570 nm in a spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Data are presented as percentage of untreated control group.
Western blot analysis
Cell culture, adenine treatment and LPS stimulations were done as explained above. Cells were collected, washed with icecold PBS, and then lysed in ice-cold lysis buffer for 30 min on ice. Lysates were centrifuged at 12,000 rpm at 4℃ for 20 min and equal amount of protein were mixed with 4× sample buffer, boiled for 5 min, subjected to SDS-PAGE and then transferred on PVDF membrane. The membrane was blocked in 5% skim milk in Tris-buffered saline containing 0.1% Tween 20 (TBS-T) for 1 h at room temperature and then incubated overnight with primary antibodies at 4℃. After washing the membrane with TBS-T, immune-reactive proteins were detected with horseradish peroxidase-conjugated secondary antibody using chemiluminescence kit. ImageJ software was used for densitometry analysis.
Preparation of cytosolic and nuclear extraction
Nuclear extracts of RAW264.7 cells were prepared using NEPER nuclear and cytoplasmic extraction reagent according to the manufacturer's instructions and supplemented with a set of protease inhibitors. Equal amount of cytosolic or nuclear protein extract was denatured in sample buffer and western blotting was done as explained above.
Determination of cellular AMP level
AMP level was measured using AMP-Glo assay kit as per the manufacturers' protocol. Briefly, RAW264.7 cells grown overnight in complete growth medium were treated with various doses of adenine for different time periods. Whole cell lysate were prepared after washing the cells with ice-cold PBS. AMP-Glo reagent and AMP detection solution were mixed according to the product manual and after a luciferase reactions, a luminescent signal was measured with a luminometer. A standard curve was generated using AMP standards in triplicate. Data were calculated and presented after normalizing the values with equal amount of protein from each sample.
Preparation of bone marrow derived mast cells (BMMCs)
BMMCs were prepared as explained elsewhere [14]. Briefly, femurs of 6 weeks old BALB/c mouse were taken and bone marrow was flushed, suspended and cultured in RPMI 1640 medium containing 10% heat inactivated FBS, 1% penicillin and streptomycin, 2 mM glutamine, 50 µM 2-mercaptoethanol, 25 mM HEPES [pH 7.4] supplemented with 10 ng/ml IL-3 in 5% CO2 incubator at 37℃. The non-adherent cells were re-seeded in the fresh medium twice a week. By 4–5 weeks in culture, the cells were used for further experiments after verifying the matured mast cells by staining with toluidine blue.
For activation experiments, BMMCs were transferred to 12-well cluster plates (2×106 cells per ml per well). Adenine was pretreated for 30 min in complete growth medium. The cells were stimulated with LPS for 6 h to measure the concentrations of TNF-α and IL-6 in the supernatants whereas stimulated for 30 min for the western blot analysis experiments.
Induction of LPS mediated acute peritonitis
The female BALB/c (6–8 weeks old) mice purchased from DBL Korea were kept and maintained under specific pathogen-free conditions according to the guidelines of the ‘Institutional Animal Care and Use Committee’ of Chungnam National University. Mice were intraperitoneally injected with adenine (10 mg/kg) or PBS and LPS (1 mg/kg) was injected after 1 h. After waiting for 1 h peritoneal lavage was collected by injecting PBS into peritoneal cavity. Lavage was cleared of any cells by centrifuging in cold condition. Amounts of cytokines were measured using ELISA kits according to the manufacturer's protocol.
RESULTS
Adenine decreases LPS-induced production of inflammatory cytokines in macrophages
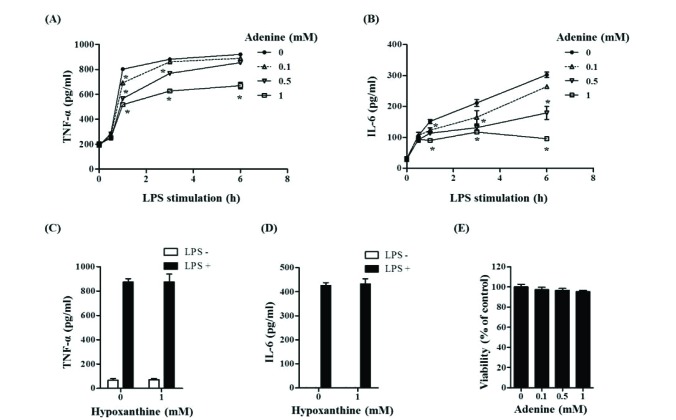
Macrophages play critical roles in innate immune reaction by producing inflammatory cytokines through activation of TLR4 on cell surfaces by LPS [29]. The effect of adenine on cytokine production was studied in LPS-stimulated RAW264.7 cells, a mouse macrophage cell line. TNF-α secretion was maximal at 3 h after LPS stimulation while IL-6 secretion peaked after 6 h (Figs. 1A and B). Treatment of adenine reduced productions of these cytokines in a dose dependent manner. One mM adenine inhibited releases of TNF-α and IL-6 by 40% and 65%, respectively (Figs. 1A and B). To know if these inhibitions are adenine specific, a structural analog of adenine, hypoxanthine, was exploited. Hypoxanthine did not impede TNF-α and IL-6 production in the LPS-stimulated cells (Figs. 1C and D). We examined whether the experimental condition applied for this study is deleterious to cells because there is a report that adenine is cytotoxic to lymphoblast cells [12]. In the experimental condition applied for this study, adenine and hypoxanthine had no effect on cell viability (Fig. 1E). These results show adenine specifically down-regulate LPS-induced production of TNF-α and IL-6 in macrophages.
Adenine inhibits LPS-induced eicosanoids production in macrophages
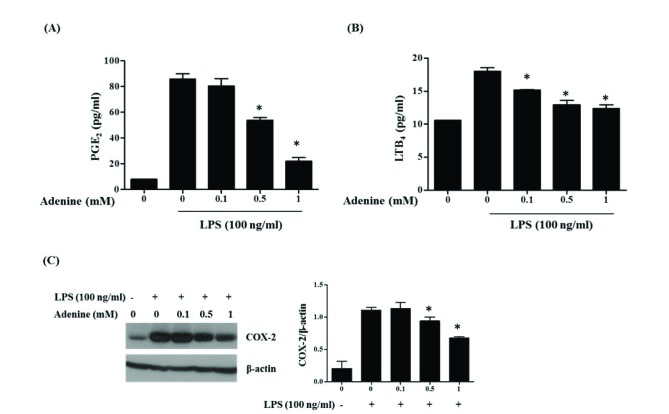
LPS stimulation to macrophages causes release of inflammatory arachidonic acid metabolites (eicosanoids) such as prostaglandins and leukotrienes made from arachidonic acid by cyclooxygenase (COX) and lipoxygenase, respectively [30]. So effect of adenine on LPS-induced eicosanoids production in RAW264.7 cells was studied. The macrophages stimulated with LPS (100 ng/ml) secreted eicosanoids PGE2 and LTB4 to the culture medium (Fig. 2). Adenine treatment inhibited the release of those eicosanoids in a dose dependent manner. One mM adenine inhibited release of PGE2 and LTB4 by 80% and 75%, respectively (Figs. 2A and B). The inducible enzyme COX-2 responding to prostaglandin synthesis was also examined. While the increase of COX-2 protein was observed 6 h after LPS treatment, this increment was inhibited by adenine treatment in a dose dependent manner (Fig. 2C). These results suggest adenine inhibit LPS-mediated synthesis of eicosanoids.
Adenine suppresses the LPS-induced activation and nuclear translocation of NF-κB in macrophages
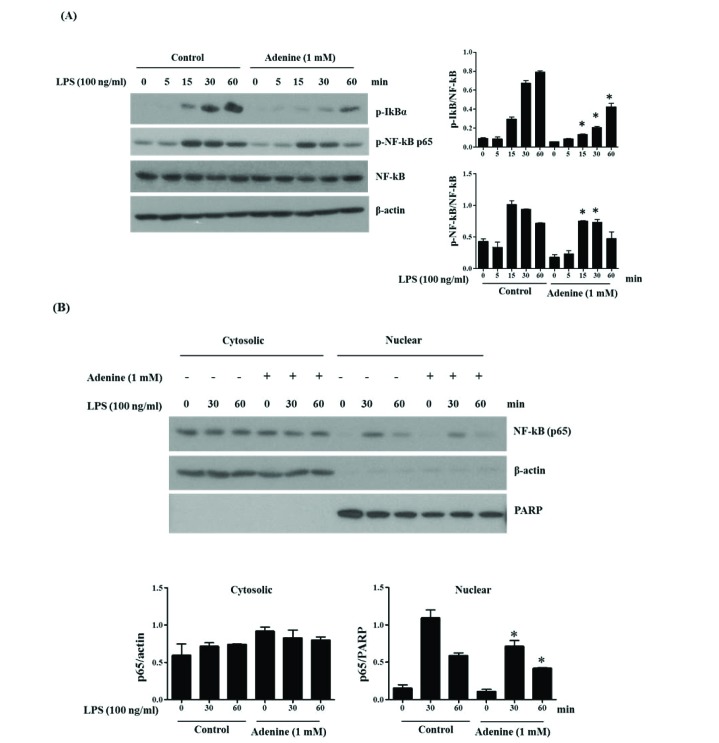
NF-κB is a transcription factor regulating the cytokine production in LPS-stimulated conditions. Activation of NF-κB occurs after the phosphorylation of IκB and subsequent nuclear translocation of NF-κB is important in activation of this signaling pathway [31]. Thus, we investigated the adenine effects on the phosphorylation and nuclear translocation of NF-κB (p65) by western blotting on LPS-induced RAW264.7 cells. Phosphorylation of IκB as well as NF-κB (p65) shows activation of these signaling starting as early as 15 min after LPS stimulation (Fig. 3). Phosphorylation of IκBα after LPS stimulation was increasing till 60 min whereas phosphorylation of NF-κB was at peak at 15–30 min. Adenine (1 mM) pre-treatment reduced the phosphorylation of IκBα and NF-κB (Fig. 3A). Also, adenine inhibited translocation of NF-κB (p65) to nucleus (Fig. 3B). These results reveal that adenine inhibits the synthesis of the inflammatory cytokines by suppressing the phosphorylation of IκB and NF-κB as well as nuclear translocation of NF-κB in LPS-stimulated RAW264.7 cells.
Adenine suppresses LPS-induced activation of MAPKs in macrophages
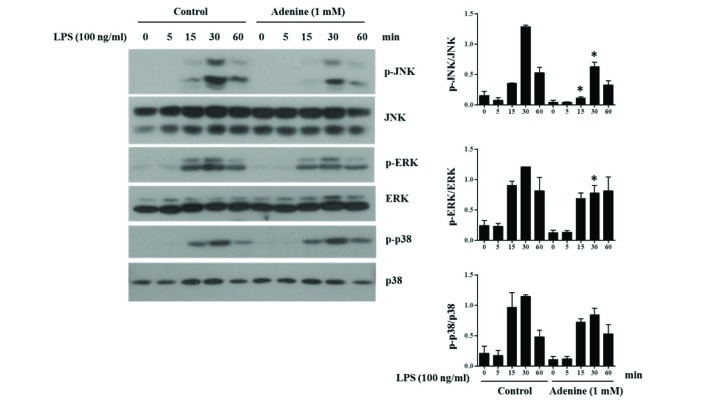
MAPKs including ERK, JNK and p38 also play essential roles in regulation of pro-inflammatory cytokines production in LPS-mediated reactions [22]. To investigate whether adenine regulates the MAPK signaling, phosphorylations of ERK, p38 and JNK were examined by western blotting. Activating phosphorylation of MAPK, ERK, p38 and JNK was detected after 15 min of LPS stimulation showing maximum intensity at 30 min. Adenine pretreatment reduced the phosphorylation of JNK and ERK whereas p38 was not affected (Fig. 4).
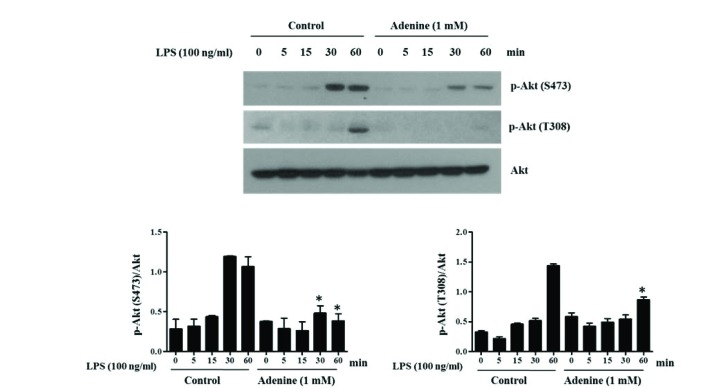
Adenine inhibits activation of Akt in LPS-induced macrophages
TLR4 mediated PI3K/Akt signaling axis is another important signaling pathway regulating immune functions [24]. So the activating phosphorylation of Akt in RAW264.7 cells after LPS stimulation was examined by western blot analysis. LPS (100 ng/ml) stimulation led to phosphorylation of Akt (Ser473 and Thr308), and adenine (1 mM) pre-treatment inhibited those phosphorylation (Fig. 5).
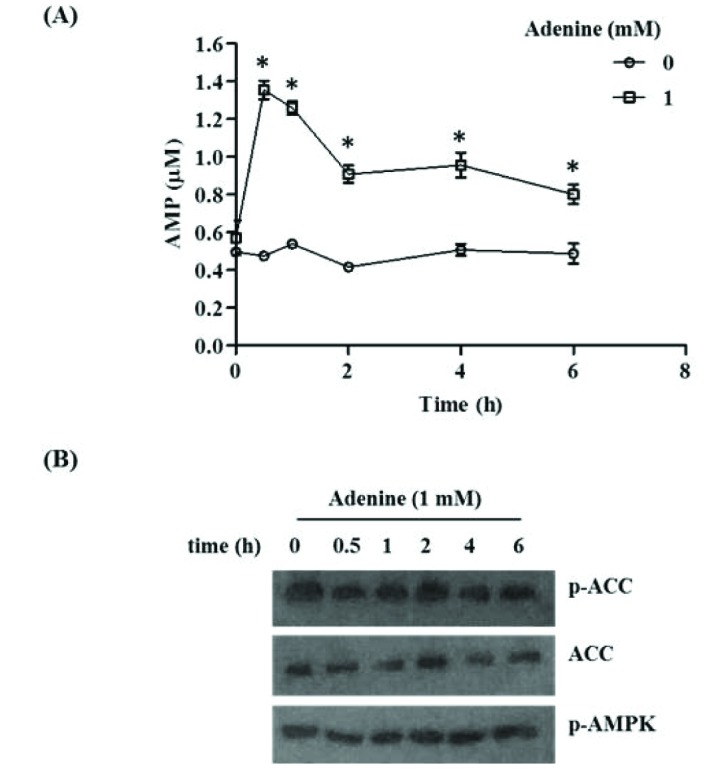
Adenine has no effect on AMPK phosphorylation in macrophages
Adenine can be irreversibly converted into AMP which in turn could lead to activation of AMPK which modulates inflammatory responses [32]. AMPK is activated by both AMP and phosphorylation on Thr172 of α subunit by upstream kinases. Previous finding has suggested that adenine activates AMPK thereby inhibiting TNF-α signaling in HUVECs and glucose uptake in NIH-3T3 cells [3233]. Hence we examined the effect of adenine on cellular AMP level and AMPK phosphorylation in RAW264.7 cells. Cellular AMP level rose after adenine treatment peaking after 30 min and gradually decreased thereafter (Fig. 6A). Adenine treatment however did not affect the activating phosphorylation of AMPK at Thr172 (Fig. 6B). Similarly, there was no effect on phosphorylation of acetyl-CoA carboxylase (ACC) at Ser79, a downstream target of AMPK (Fig. 6B). These results suggest that although adenine increases AMP amounts, it is not involved in AMPK activation.
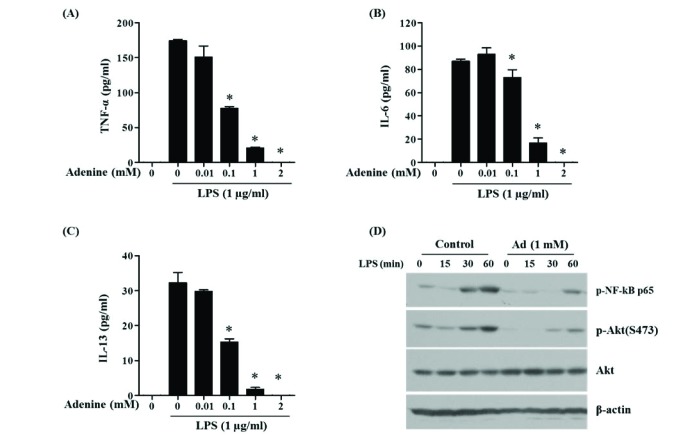
Adenine inhibits LPS-induced activation of mast cells
The anti-inflammatory effect of adenine on LPS-stimulated macrophage demonstrated above was also examined in mast cells. They respond to LPS via TLR4 in host defense against Gram negative bacteria, in addition to their role in allergic diseases [28]. Secretion of inflammatory cytokines TNF-α, IL-6 and IL-13 in LPS-stimulated primary mast cells BMMCs was inhibited by adenine treatment in a dose dependent manner (Figs. 7A–C). Signaling molecules involved in TLR4-mediated cytokine synthesis, NF-κB (p65) and Akt (Ser473), were activated by LPS and these responses were attenuated by adenine (Fig. 7D). These results reveal that adenine hinders LPS-induced inflammatory responses of macrophages and mast cells.
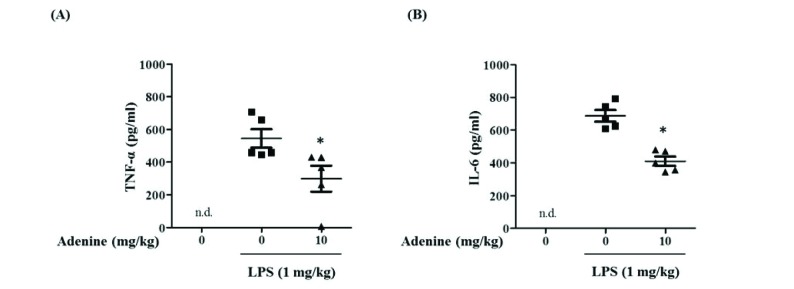
Adenine inhibits cytokine production in LPS-induced acute peritonitis
Anti-inflammatory role of adenine in cultured cells were examined in mice. After injection of LPS (1 mg/kg) to peritoneal cavities of mice, peritoneal lavages were collected to measure levels of cytokines. LPS injection caused production of TNF-α and IL-6 in peritoneal cavities and adenine (10 mg/kg) inhibited the production of TNF-α and IL-6 cytokines (Figs. 8A and B). These results demonstrate that adenine attenuates inflammatory responses of macrophages, primary mast cells and peritoneal cells in mice.
DISCUSSION
Besides being a structural component of biomolecules, there is a rising trend to study adenine as a modulator of several pathophysiological conditions such as allergy and inflammation. In this study, we show that adenine inhibits LPS-induced inflammation in macrophage cells, primary mast cells and peritoneal cells in mice. It attenuates release of inflammatory cytokines (TNF-α, IL-6) and lipid mediators (PGE2, LTB4) in macrophages and mast cells by inhibiting NF-κB, MAPK and PI3K/Akt signaling. It also inhibits LPS-induced secretion of the cytokines in peritoneal cavity of mice.
Adenine inhibited LPS-stimulated secretion of inflammatory cytokines TNF-α and IL-6 from macrophages (Figs. 1A and B); TNF-α, IL-6 and IL-13 from mast cells (Figs. 7A–C); TNF-α and IL-6 from peritoneal cells in mice (Fig. 8). They mediate many physiological effects such as septic shock, inflammation, cytotoxicity and cellular metabolism [3435]. Inhibition of secretion and action of the cytokines is important in treatment of inflammatory conditions. In addition to the role in allergy, mast cells play crucial roles in defense against bacteria as they can respond against LPS through TLR4 [2735]. Peritoneal cells in mice including macrophages and mast cells play crucial role in LPS-mediated inflammatory response [3637]. From inhibitory action of adenine to secretion of inflammatory cytokines by three kinds of LPS-stimulated cells, we suggest that adenine is an immune suppressor attenuating LPS-mediated pathology.
Adenine prevented LPS-mediated activation of NF-κB (Figs. 3A and 7D), MAPKs (Fig. 4) and Akt (Figs. 5 and 7D) which are important for production of inflammatory cytokines. Inhibition of this signaling has important therapeutic potential for inflammation and cancer [38]. LPS-stimulated TLR4s are dimerized and recruit different adapter molecules such as MyD88, leading to activation of NF-κB, MAPKs and Akt [202223]. The activated NF-κB translocate from cytosol to nucleus and regulates transcription of many genes encoding cytokines, chemokines, nitric oxide synthase, COX-2 and pro-inflammatory proteins [39]. ERK regulates the activity of cPLA2 which produces arachidonic acid, the precursor of prostaglandins and leukotrienes. Also NF-κB and MAPK both regulate COX-2 thereby affect the production of arachidonic acid metabolites [2239] so the effect of adenine on eicosanoid production might come through inhibition of NF-κB and MAPK mediated signaling. Akt stimulates the transactivation potential of the RelA/p65 subunit of NF-κB and activation of NF-κB [2440]. Similar to our results, adenine reduced the JNK and IκB phosphorylation in TNF-α induced activation of human intestinal epithelial cells (Caco-2) [15]. These results show that adenine suppresses LPS-mediated TLR4 signaling through inhibition of NF-κB, MAPK and Akt signaling.
Adenine is cell-permeable and can be converted to AMP in cells by an enzyme APRT [5]. The AMP thereby activates AMPK to promote various physiological effects. Several reports suggest that adenine could be a novel and simplest AMPK activator [3233]. However, in our experimental settings, we could not detect significant change in phosphorylation of AMPK at Thr172 and its downstream target ACC after adenine treatment (Fig. 6B) although cellular AMP level was increased (Fig. 6A). These results suggest that AMP is necessary but not sufficient for AMPK activation in certain conditions. There are similar reports with our observation. AICAR is metabolized to ZMP in cells that is the functional and structural analog of AMP in terms of AMPK activation. It has anti-inflammatory effects in LPS-stimulated macrophages in an AMPK independent manner [4142]. Therefore we propose that the anti-inflammatory effect of adenine is independent on AMPK activation although it increases cellular AMP level. The action mechanism of adenine in LPS-mediated signaling remains to be solved.
Adenine at 1 mM, which had no cytotoxic effect, was used to get significant effect in this study. Similarly adenine is known to inhibit allergic response [14] as well as immunological functions of lymphocytes at pharmacological concentration in millimolar (mM) scale [1]. Its plasma concentration is 70 nM in healthy subjects while 1450 nM in patients with chronic renal failure [43]. However, the adenine concentration in local tissues may reach to mM level after immune stimulation or in pathologic conditions because these circumstances make immune cells proliferate and then produce adenine substantially [5].
In conclusion, we reveal that adenine inhibits the LPS-induced inflammatory reactions via inhibition of NF-κB, MAPK and PI3K/Akt pathways. Our results provide new insight into adenine as a potential therapeutic agent to inflammatory diseases.
 XML Download
XML Download