

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Muscle atrophy, caused by a decrease in physical activity or aging, leads to problems related to an increase in body fat and decreased muscle strength and cardiovascular endurance, resulting in fatigue [1234] and deterioration of bone health [56], ultimately affecting the activities of daily living and quality of life [789]. Thus, efforts to improve muscle atrophy in the increasing elderly population should be explored.
Hypertrophy of the skeletal muscles is required to prevent muscle atrophy [10], and exercising with weights, including resistance exercise training, is effective [11]. According to recent studies, even though muscle size and strength can be increased with low-intensity resistance training [12], higher-intensity resistance training (i.e., 80% of one repetition maximum) is more effective [131415]. However, many adults, including elderly people (more than 70%), are inactive or sedentary, mainly participating only in light physical activities such as stretching or walking [161718], making them at increased risk of various injuries, such as muscle damage, compared with young adults [1920]. Therefore, exercising with the lowest intensity possible may be helpful in reducing the risk of injuries. However, as the intensity of the exercise decreases, its effects inevitably decrease. Hence, we hypothesized that if the effects of high-intensity exercise can be generated at a lower intensity using exercise mimetics, health could be safely maintained without the risk of injuries.
Ursolic acid (UA), a pentacyclic triterpenoid compound that is found in herbs and fruits, has various biological activities [212223]. UA is reported to improve the insulin/insulin-like growth factor 1 (IGF-1) signaling pathway; reduce the expression of muscle ring-finger protein 1 (MuRF1) and atrogin-1, a muscle-specific F-box protein [24]; and induce muscle hypertrophy. It has effects similar to those of resistance training [1242526]. In addition, UA is considered to be an effective compound for the treatment of obesity and diseases related to obesity, by decreasing high fat diet-induced obesity and insulin resistance [27]. UA is considered to be an exercise mimetic that is effective for the prevention of, and improvement in, various diseases caused by muscle strength reduction and body fat increase. Although UA intake has a positive effect on muscle atrophy and various disease management by muscle atrophy, it is difficult to improve all fitness factors, including flexibility, balance, and coordination. Furthermore, it does not have a positive effect on emotional health such as pleasure and happiness, and the effects of many nutrients are somewhat controversial. UA also has cell death-inducing properties [2829], anti-inflammatory and pro-inflammatory properties [30], and pro-angiogenic and anti-angiogenic properties [3132]. Therefore, strategies for minimizing the negative effects (such as lowering the UA dose) and maximizing the positive effects are needed. However, the positive effect of UA may not be fully realized in this case. Thus, we proposed the following hypothesis. A low concentration of UA can help improve several health-related disadvantages associated with muscle atrophy if it can promote low risk of injury with effective yet low-intensity exercise effects.
This study was aimed at evaluating the use of UA as an exercise mimetic to safely and effectively improve muscle atrophy-related physical dysfunction. We used an animal model of skeletal muscle atrophy, one of the major characteristics of aging, and analyzed the concurrent treatment effects of UA and low-intensity treadmill exercise to determine any benefits related to improving muscle atrophy.
Go to : 
METHODS
Animals and experimental design
We used 8-week-old male Sprague-Dawley (SD) rats (n=40), weighing approximately 250 g (Central Lab, Animal Inc., Seoul, Korea). All the SD rats were allowed to acclimate for 1 week. The rats were randomly allocated into four groups: SED, sedentary; UA, daily intraperitoneal UA injection, 5 mg/kg; EX, low-intensity (10–12 m/min, 0° grade) treadmill exercise; UEX, daily intraperitoneal UA injection, 5 mg/kg, and low-intensity (10–12 m/min, 0° grade) treadmill exercise. Rats were given ad libitum water and food (carbohydrate, 58.9%; fat, 12.4%; protein, 28.7%; Purina Corp.MO, USA) during the experimental period. Animals were housed in an environmentally controlled laboratory (temperature, 21℃; humidity, 40%–60%) with a 12:12-h dark and light cycle.
The left hind limb of each animal was immobilized using the method employed by Coutinho et al. [33], and muscle atrophy was induced over 10 days. After 10 days, the animals were treated with UA, EX, and UA plus EX for 8 weeks. Body weight was measured once per week during the 8-week period. After 8 weeks of treatment, the rats were anesthetized using sodium pentobarbital (50 mg/kg of body weight). Three rats per group were used for the micro-computed tomography (micro-CT) scan of both hind limbs for 3D structural analysis. Hind limbs were removed, fixed, and preserved using 10% paraformaldehyde. In the remaining rats (7 rats per group), epididymal and retroperitoneal fat 24 h after the last training session was extracted. In addition, tibialis anterior (TA), soleus (SOL), and gastrocnemius (GAS) muscles were removed from both hind limbs, with the contralateral, nonimmobilized leg being used as an internal control. The tissues were weighed using an electronic scale (EPG213; Ohaus Co., NJ, USA). The present study received approval from the Institutional Animal Care and Use Committee in Chonbuk National University (IACUC approval no. CBNU-2017-0008).
Unilateral immobilization
Rats were anesthetized with pentobarbital sodium (50 mg/kg of body mass; Fort Dodge Animal Health, USA). The left knee joint was stretched, and the ankle joint was fixed by casting in a neutral posture [3435]. During the immobilization, the rats were monitored for respiration state, discomfort, hind limb edema, and skin color changes. The animals were allowed to move freely, preventing movement of the immobilized leg alone. The muscles from the contralateral non-immobilized leg were used as controls in all experiments.
Ursolic acid injection and exercise program
UA was administered by intraperitoneal injection (5 mg/kg) once per day, after it was dissolved in distilled water containing 0.1% Tween 80 (Sigma, St. Louis, MO, USA) [36]. For all other groups, the same amount of vehicle (distilled water containing 0.1% Tween 80) was intraperitoneally injected. Intraperitoneal administration is a commonly used method in animal studies and can be a practical alternative in the late phase, but the initial uptake is lower than that of intravenous and oral routes [37].
The low-intensity exercise protocol was based on partial revisions to the methods by Kondo et al. [38]. The low-intensity exercise was carried out for 8 weeks using a treadmill (Omnipacer model LC-4, Omnitech, USA). Before the exercise, training for acclimatization on a treadmill was conducted for 3 days, 15 min per day, at a speed of 5–10 m/min on a 0° gradient. For the experiment, rats walk or run on the treadmill for 30 min/day 3 days/week for 8 weeks. The gradient of treadmill was set to 0°, and the speed was set at 10 m/min for the first 3 weeks, 12 m/min for the next 3 weeks, and then 15 m/min (45–55% Vo2max) [39] for the last 2 weeks.
Micro-computed tomography and three-dimensional structural analysis
Scanning was carried out using a SkyScan 1076 system (Bruker microCT; Kontich, Belgium) at the following settings: 12.25-µm resolution, 70 kV, 141 µA, and Al 0.5-mm filter. The images were reconstructed using NRecon software (Bruker microCT; Kontich). The cross-sectional images of the hind limb were captured using CTAn and CTvox software (Bruker microCT; Kontich).
Quantitative real-time polymerase chain reaction
After total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) from the GAS, it was purified using the DNA-free™ Kit (Ambion, CA, USA). After the concentration of purified RNA was measured using an ultraviolet spectrophotometer (UV-mini 1240, Shimazu Co., Japan), reverse transcription was conducted using 0.5 µg of RNA using the iScript™ cDNA Synthesis Kit (Bio-Rad, CA, USA). Real-time polymerase chain reaction analysis was performed with the produced cDNA as a template using the iQ5 Multicolor Real-Time PCR Detection System (Bio-Rad, CA, USA). This was achieved by mixing with MuRF1 (sense: TGTTCTGGTAGGTCGTTTCCG, antisense: ATGCCGGTCCATGATCACTT), Atrogin-1 (sequence: CCATCAGGAGAAGTGGATCTATGTT, antisense: GCTTCCCCCAAAGTGCAGTA), Akt (sense: GTGCTGGAGGACAATGACTACGG, antisense: AGCAGCCCTGAAAGCAAGGA), mTOR (sense: TCCTGAAGAACATGTGCGAG, antisense: CCAAAGTACAAGCGAGAGGC) and 18S rRNA's specific primer with the reaction enzyme reagent, 2X SYBR Green Supermix (Bio-Rad, CA, USA). The mRNA expression level was quantified as the ratio to 18S rRNA.
Western blot analysis
Muscle extracts were prepared and Western blotting was performed as described previously [40]. Blots were probed with the following antibodies: Akt, P-Akt, mTOR, P-mTOR, MuRF-1, Atrogin-1 (Bio-Rad, CA, USA), and β-actin (Sigma, MO, USA). The blots were then incubated with the appropriate horseradish-conjugated secondary antibodies (Jackson Immunoresearch Laboratories, PA, USA). Antibody bound protein was detected by ECL substrate (Bio-Rad, CA, USA). Band intensity was visualized and calculated using a ChemiDoc XRS+ (Bio-Rad, CA, USA).
Endurance capacity
After 8 weeks of treatment, the endurance capacity of each rat was measured using an exercise tolerance protocol on the treadmill. In line with the protocol for a progressive exercise test, treadmill exercise at a speed of 10 m/min was initially carried out at a 15° gradient for 5 min, with the speed increasing at 2 m/min each minute. Any rat that could not run anymore and stayed on the electrical shock bar (on which there was no electric current) was encouraged to continue to run by stimulating the hind limbs with high pressure air. If the rat stayed on the shock bar for more than 3 s despite the air stimulation, the test was suspended [41] and total running time and distance were calculated. The equation used by Park et al. [42] to calculate total work was used: work (J)=force [body weight (kg)]×vertical distance [sin (15°)×speed (m/min)×time (min)].
Statistical analysis
The data are presented as mean±standard error (SE). The unpaired t-test was initially conducted for comparison of muscle mass based on pre-immobilization and 10 days after, to determine the muscle atrophy level. A one-way analysis of variance analysis was carried out to verify the differences by each variable between groups. A Bonferroni post hoc test was conducted to determine the significance. The significance level was set at p<0.05. The statistical analyses were performed with SPSS 18.0 (SPSS Inc., Chicago, IL, USA).
Go to :
RESULTS
Hind limb immobilization induces muscle atrophy
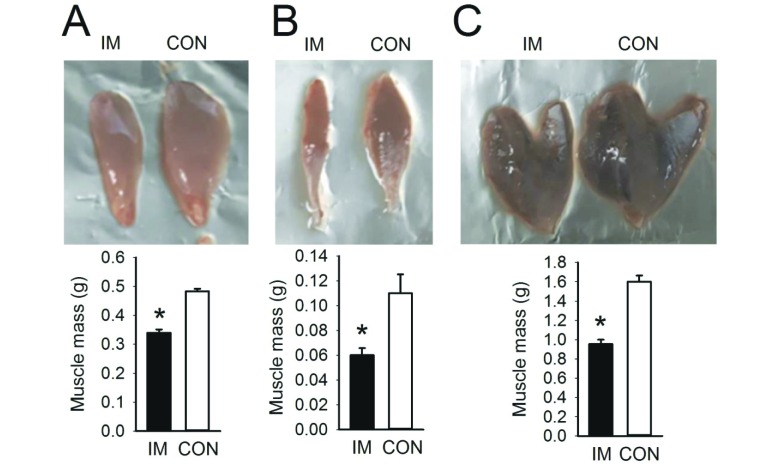
A muscle atrophy model was generated to identify the effects of low-intensity treadmill exercise and UA intake on various physical dysfunctions associated with muscle atrophy. The effect of hind limb immobilization for 10 days was evaluated to determine the extent of muscle atrophy. It was confirmed that the muscle mass in the hind limbs was significantly reduced by immobilization, as shown in Fig. 1.
Concurrent treatment with low-intensity treadmill exercise and UA reduces body weight and visceral fat
The effects of low-intensity treadmill exercise and UA intake for 8 weeks on body weight and visceral fat were evaluated (Fig. 2). Although 8 weeks of low-intensity treadmill exercise and UA treatment had an inadequate effect on body weight, the body weight of the UEX group was significantly lower than the SED and UA groups (p<0.05). Epididymal fat was significantly lower in the EX and UEX groups compared with the SED group (p<0.05), and retroperitoneal fat was significantly lower in the UEX group compared with the SED group (p<0.05).
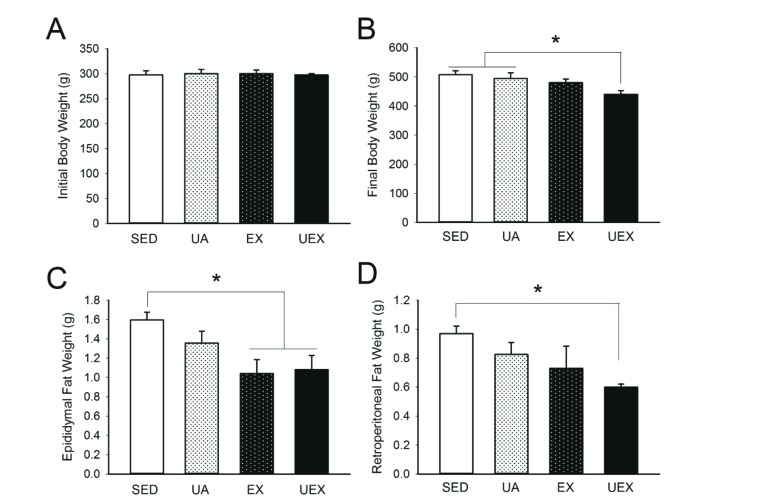 | Fig. 2Reduction in final body weight and fat mass through UA supplementation with exercise.Rats were given UA and/or performed treadmill exercise for 8 weeks. Data are presented as mean±SE. (A, B) Initial and final body weights were measured before immobilization and before sacrifice (n=7 rats per group). (C, D) Weight of epididymal and retroperitoneal fat pads (n=6 rats per group). SED, sedentary; UA, ursolic acid; EX, exercise; UEX, ursolic acid plus exercise. *p<0.05, one-way analysis of variance.
|
Concurrent low-intensity treadmill exercise and UA treatment reduces immobilization-induced muscle atrophy
Fig. 3 shows the results as a ratio to the contralateral muscle of the improvement of the muscle mass according to the treatment for 8 weeks. All the muscles analyze tended to improve according to treatment. Specifically, the UEX group was significantly improved TA and GAS muscles compared to SED group (p<0.05) (Fig. 3A). The same result was shown in terms of cross-sectional area by micro-CT analysis (Fig. 3B). There was no significant difference between the immobilization-induced muscle and contralateral muscle of the UEX group compared with the other groups.
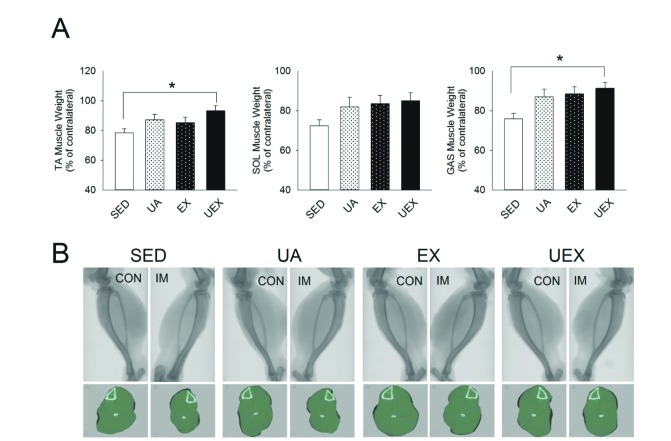 | Fig. 3Improvement in atrophied hind limb muscle mass through UA supplementation with treadmill exercise.(A) Skeletal muscle atrophy of the TA, SOL, and GAS induced by IM is expressed in percentage of difference from the contralateral CON leg (non-immobilized). Data are presented as mean±SE. *p<0.05, one-way analysis of variance. (B) Micro-computed tomography (CT) scans of a rat hind limb. Lateral view of a whole limb (top) and mid-tibial transverse micro-CT scans of the same rat hind limb (bottom). TA, tibialis anterior; SOL, soleus; GAS, gastrocnemius; IM, immobilization; CON, control; SED, sedentary; UA, ursolic acid; EX, exercise; UEX, ursolic acid plus exercise.
|
Concurrent low-intensity treadmill exercise and UA treatment reduces atrophic gene expression
We analyzed muscle atrophy- and hypertrophy-related gene expression to evaluate how the concurrent low-intensity treadmill exercise and UA treatment were effective in improving skeletal muscle mass. As a result, reduction of the mRNA and protein expression of atrophy-related genes was confirmed (Figs. 4A and 4C). Significant differences from the SED group were observed in the UEX group in terms of MuRF1, and in the UA and UEX groups in terms of Atrogin-1 (Fig. 4A). The content of MuRF1 and Atrogin-1 protein was significantly differences between SED group and UEX group. However, mRNA and protein expression of Akt and mTOR, the genes related to muscle hypertrophy, was not different between the groups.
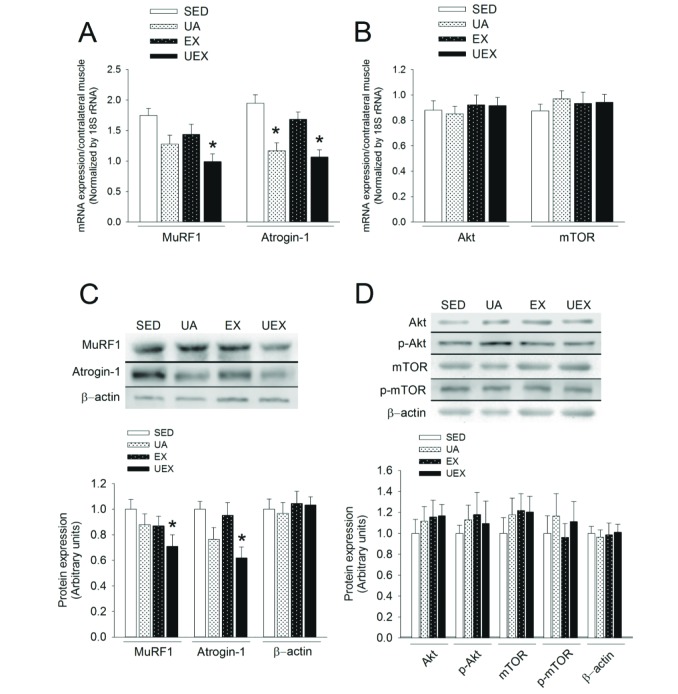 | Fig. 4Reduction in atrophic gene expression through concurrent low-intensity treadmill exercise and UA treatment.Relative mRNA expression levels of atrophy- (A) and hypertrophy- (B) related genes and the content of atrophy- (C) and hypertrophy- (D) related protein after 8 weeks of treatment in the gastrocnemius muscle (n=6 rats per group). Signals were quantified and normalized against the contralateral CON leg (non-immobilized). Data are presented as mean±SE. *p<0.05, one-way analysis of variance. CON, control; SED, sedentary; UA, ursolic acid; EX, exercise; UEX, ursolic acid plus exercise.
|
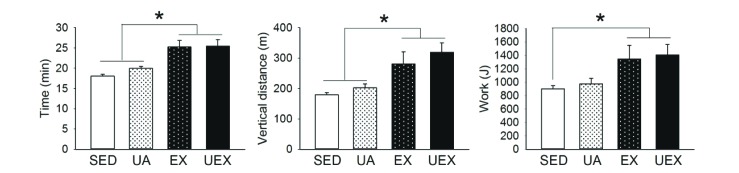
Endurance capacity improves with low-intensity treadmill exercise
As determined by performing an endurance capacity test after 8 weeks of treatment, the exercise time, running distance, and total work in the EX and UEX groups significantly improved compared with the SED group (Fig. 5). However, UA did not have an additive effect in endurance capacity at all.
Go to :
DISCUSSION
This study was conducted to evaluate the effects of UA and low-intensity treadmill exercise on the improvement of muscle atrophy and exercise capacity caused by physical dysfunctions such as aging. Based on these results, we tried to confirm the possibility of UA as an exercise mimetic for the safe and effective improvement of physical dysfunctions. Exercise mimetics are defined as biologically active compounds that have an effect on exercise by activating signaling pathways similar to exercise [43]. Interest in exercise mimetics continuously increased after Narkar et al. [44] reported that the AMPK-PPARδ pathway not only can improve adaptation according to training, but medicine can be used to target an increase in endurance without performing exercise [44454647]. However, most relevant research has focused on identifying exercise mimetics for treatment of obesity through endurance capacity improvement or to treat metabolic disorders [4445464748]. UA was reported to affect obesity and improve insulin resistance [27] and improve muscle hypertrophy or muscle atrophy [1242526] through increased endurance capacity. This study confirmed that concurrent low-intensity exercise and UA treatment can generate an effect comparable to that of high-intensity exercise and analyzed the possibility of UA as an exercise mimetic. Hence, a muscle atrophy model was made based on casting-induced hind limb immobilization in SD rats. The hind limb immobilization used in this study for 10 days reduced 30%–45% of the muscle volume and thus confirmed that hind limb immobilization could be used as a model for muscle atrophy (Fig. 1). This study analyzed the effects of 8 weeks of treatment using this model.
Muscle atrophy is closely related to physical activity reduction [49], and weight gain as a result of the reduction in physical activity causes various health problems [50]. Therefore, preventative treatment is required. According to the results of the present study, 8 weeks of UA treatment and low-intensity treadmill exercise did not significantly contribute to weight gain prevention (Fig. 2B). However, low-intensity treadmill exercise resulted in some visceral fat reduction by reducing (p<0.05) epididymal fat. The retroperitoneal fat weight was decreased 25% but no significantly (p=0.068) changed in EX group. Both epidydimal and retroperitoneal fat were reduced in the UEX group (Figs. 2C and 2D). We have not determined the reason for the different responses in epididymal fat and retroperitoneal fat in the EX and UEX groups, but we noted a decrease in visceral fat in the EX group. This result indicates that weight gain was effectively prevented with the combined treatment. UA reduces interscapular fat by increasing body temperature and energy expenditure with increasing uncoupling protein (UCP) 1 in brown adipose tissue [27]. In addition, it increases energy expenditure by increasing the UCP 3/AMPK-dependent pathway [51] as well as enhancing β-oxidation in skeletal muscle [5152]. The effects of UA are similar to the increase of fat oxidation in skeletal muscle [535455] by activating carnitine palmitoyl transferase (CPT1) by controlling the synthesis of malonyl CoA by the fatty acid's increased oxidation through exercise, namely control of acetyl CoA carboxylase activity by increased AMPK activity. Therefore, it is possible that UA may be an exercise mimetic that could promote weight loss, but there are limitations in preventing weight gain from muscle atrophy through UA alone. Hence, concurrent low-intensity endurance training and UA intake can be a very effective treatment method.
UA decreases muscle atrophy by decreasing the expression of atrophy-related genes and increases the expression of hypertrophy-associated genes, inducing muscle hypertrophy [2426]. It decreases the expression of MuRF1 and Atrogin-1 and improves the insulin-like growth factor 1 (IGF-1) signaling pathway for muscle hypertrophy. Resistance training also increases muscle hypertrophy through increased mTOR signaling action [5657] and decreases muscle atrophy-related gene expression [958]. In this regard, although UA is considered to be an exercise mimetic for muscle atrophy improvement, this study could not confirm the effect of UA alone, which may be similar to the weight loss effect induced through exercise. However, UA administration tended to increase the mass of all muscle compared with the SED group but does not significant (Fig. 3). This may be related to the concentration of UA. A study of 200 mg/kg of UA treatment showed that muscle atrophy was reduced, muscle hypertrophy was stimulated [24], as well as a potential candidate for treatment of pathological conditions associated with muscle atrophy [59]. However, in this study, UA was treated at a concentration of 5 mg/kg. This concentration improved the soleus muscle mass in the sciatic nerve injury animal model [36], but is much lower than the 200 mg/kg of concentration. Therefore, it would be necessary to carry out a study to compare the different concentrations. The EX group also tend to increase the mass of all muscle compared with the SED group but no significant (Fig. 3). Endurance training is known to improve muscle function, neuromuscular junction, metabolism, and motor-unit adaptability [60]. Moreover, physical activity plays a role in delaying muscle mass loss [60]. Souza et al. [61] report that endurance exercise training prevents heart failure-induced skeletal muscle atrophy. Additionally, Ji and Kang [62] suggest that PGC-1a, which plays an important role in mitochondrial biosynthesis in skeletal muscle, may be contributed to improve the quality of life of elderly people by increasing skeletal muscle health such as suppression of muscle atrophy. Although resistance training is generally associated with muscle hypertrophy, this study confirmed that muscle atrophy can be effectively improved with low-intensity endurance exercise with UA intake (Fig. 3). These results suggest that the effects of UA and low-intensity exercise are combined.
Atrophy due to aging is mainly caused by a decrease in both number and size of type II fibers [63]. Myosin heavy-chain isoform of GAS muscle consists of about 6%, 8%, 22%, and 60% for types I, IIa, IIx, and IIb fibers, respectively [64]. Thus, we used GAS muscle to identify signaling pathway to confirm the possibility of UA as exercise mimetics to improve physical dysfunction such as aging. UA is effective for Akt/mTORC1 signaling within the skeletal muscle that is associated with muscle hypertrophy [2765], but some studies reported no effect [6667]. Furthermore, MuRF1 and Atrogin-1 expression, which are atrophy-related genes, which are atrophy-related genes, are controlled by UA [2426]. Endurance exercise also prevents muscle atrophy to occur via a decrease in catabolic factors such as atrogin-1, MuRF1, FoxO1, and myostatin [61]. However, as shown in Fig. 4 of this study, UA and low-intensity endurance exercise did not significantly affect the mechanism associated with atrophy and hypertrophy. However, the muscle atrophy improvement following UA intake concurrent with low-intensity endurance exercise is effective because the expression of MuRF1 and Atrogin-1 in the UEX group is significantly decreased. It is considered that the effects of anti-catabolic mechanism of UA and low-intensity endurance exercise are additive to each other. Therefore, it will be necessary to clarify whether the UA and endurance exercise effects are achieved through different mechanisms or through the same mechanism and whether the effects of each other are additive.
UA increases running distance and exercise time [262768], which not only reduces serum levels of fatigue-related factors such as lactate and lactate dehydrogenase [26], but also increases PGC-1α and mitochondrial transcription factor A (mtTFA) expression, which plays a key role in mitochondrial biogenesis, mitochondrial enzymes citrate synthase activity, and COX mRNA expression [68], thereby increasing endurance exercise capacity. In the present study, only the EX and UEX groups showed an increase in exercise time, running distance, and total work compared to the SED group (Fig. 5). Low-intensity endurance exercise alone was found to improve endurance exercise capacity. However, we could not confirm the effect of UA alone on improving endurance capacity. This is also due to the difference in concentration of UA as the result of muscle mass. UA increases serum irisin in human study [25] and increases mitochondrial biogenesis through activation of AMPK-PGC-1α in C2C12 myotubes [68]. Therefore, it may be necessary to evaluate the effect of PGC-1-related mechanism in skeletal muscle according to UA concentration.
In conclusion, this study analyzed the effects of concurrent treatment with UA and low-intensity treadmill exercise to improve skeletal muscle atrophy using an animal model. The effect of UA alone on muscle atrophy could not be confirmed; however, positive effects could be confirmed for UA combined with low-intensity endurance exercise. Further studies will be needed to verify the differences in concentration of UA and need to additionally analyze the relevant signaling pathway to confirm a mechanism for use of UA as an exercise mimetic. This study is meaningful in that UA was identified to have a possible use as an exercise mimetic for reduction of body weight or visceral fat mass and inhibition of muscle atrophy.
Go to :
 XML Download
XML Download